Inhibition of K⺠outward currents by linopirdine in the cochlear outer hair cells of circling mice within the first postnatal week
- Affiliations
-
- 1Department of Physiology, College of Medicine, Dankook University, Cheonan 31116, Korea. ansil67@hanmail.net
- KMID: 2371044
- DOI: http://doi.org/10.4196/kjpp.2017.21.2.251
Abstract
- Inhibition of K⺠outward currents by linopirdine in the outer hair cells (OHCs) of circling mice (homozygous (cir/cir) mice), an animal model for human deafness (DFNB6 type), was investigated using a whole cell patch clamp technique. Littermate heterozygous (+/cir) and ICR mice of the same age (postnatal day (P) 0 -P6) were used as controls. Voltage steps from -100 mV to 40 mV elicited small inward currents (-100 mV~-70 mV) and slow rising K⺠outward currents (-60 mV ~40 mV) which activated near -50 mV in all OHCs tested. Linopirdine, a known blocker of K⺠currents activated at negative potentials (I(K,n)), did cause inhibition at varying degree (severe, moderate, mild) in K⺠outward currents of heterozygous (+/cir) or homozygous (cir/cir) mice OHCs in the concentration range between 1 and 100 µM, while it was apparent only in one ICR mice OHC out of nine OHCs at 100 µM. Although the half inhibition concentrations in heterozygous (+/cir) or homozygous (cir/cir) mice OHCs were close to those reported in I(K,n), biophysical and pharmacological properties of K⺠outward currents, such as the activation close to -50 mV, small inward currents evoked by hyperpolarizing steps and TEA sensitivity, were not in line with I(K,n) reported in other tissues. Our results show that the delayed rectifier type K⺠outward currents, which are not similar to I(K,n) with respect to biophysical and pharmacological properties, are inhibited by linopirdine in the developing (P0~P6) homozygous (cir/cir) or heterozygous (+/cir) mice OHCs.
Keyword
MeSH Terms
Figure
-
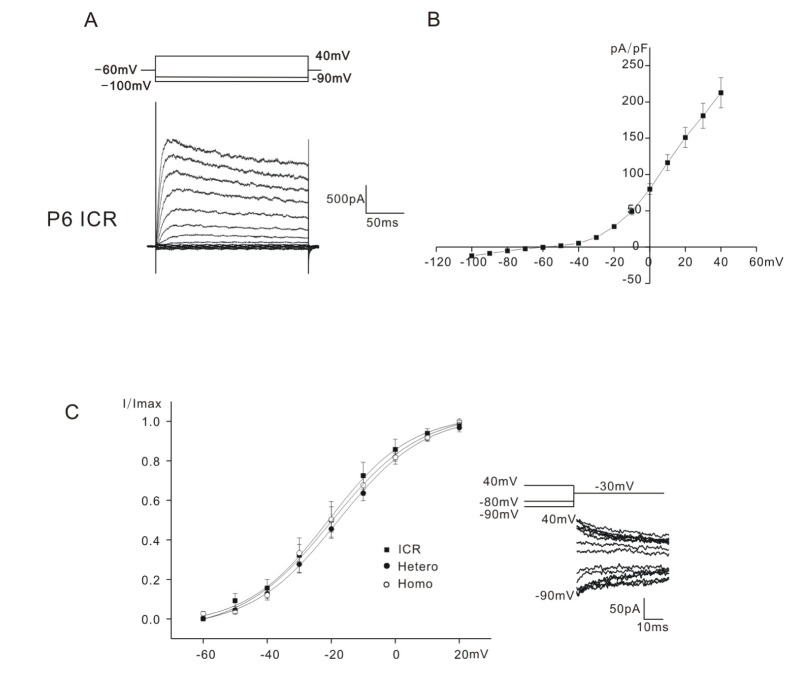
Fig. 1 Whole cell K+ currents of P0~P6 mice OHCs. Current traces recorded from P6 ICR mice is shown in A.Currents were elicited by depolarizing voltage steps from –100 mV to 40 mV with 10 mV increment (holding potential: –60 mV). Voltage protocol is shown above the current traces. Current density-voltage curve of ICR mice is shown in B. Activation curves for ICR, heterozygous (+/cir), and homozygous (cir/cir) mice are shown in C (filled square: ICR, filled circle: heterozygous (+/cir) mouse (Hetero), hollow circle: homozygous (cir/cir) mouse (Homo)). The activation curves in C were obtained by plotting the normalized tail currents at -30 mV against different prepulse potentials (–90 mV~40 mV). The voltage protocol and tail currents are shown in inset.
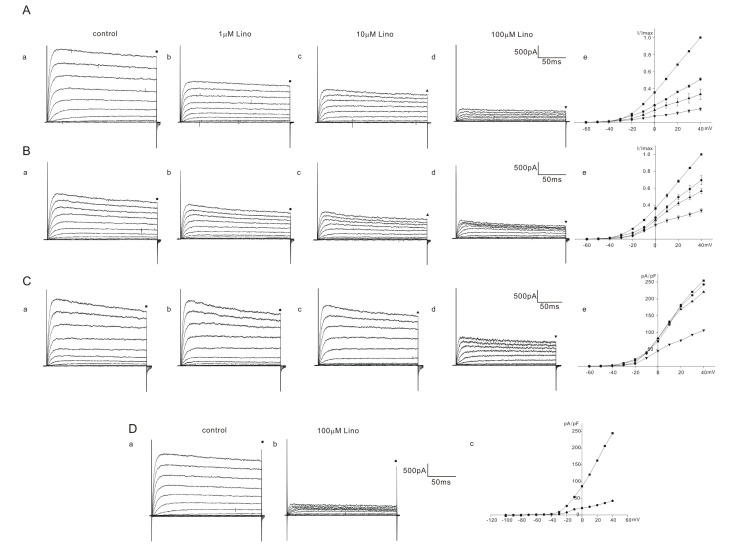
Fig. 2 Effect of linopirdine on K+ currents in homozygous (cir/cir) mice OHCs.The varying degree of inhibition of K+ currents by linopirdine (1 to 100 µM) is shown in A, B, and C. Currents were elicited by depolarizing voltage steps from –60 mV to 40 mV (the holding potential was –60 mV). Reduced currents were normalized (I/Imax) with the peak currents at 40 mV before drug application. Normalized currents – voltage curves are shown in Ae, Be, and Ce. Marks above the current traces indicate the measuring points and linopirdine concentrations. Fig. 2C shows the linopirdine effects recorded in one cell. Fig. 2D shows the linopirdine effects recorded in one cell, currents were elicited by depolarizing voltage steps from –100 mV to 40 mV (the holding potential was –60 mV). Linopirdine was not effective at hyperpolarizing voltage range (–100 mV~–60 mV).
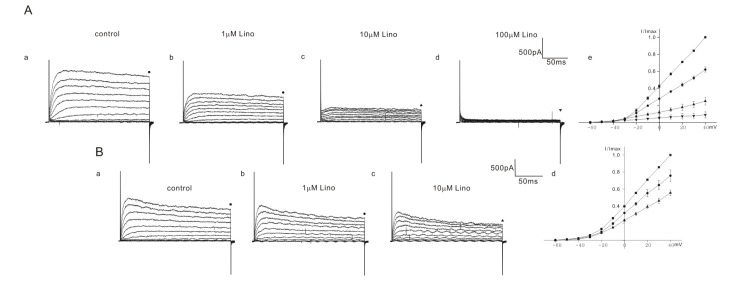
Fig. 3 Effect of linopirdine on K+ currents in heterozygous (+/cir) mice OHCs.The varying degree of inhibition of K+ currents by linopirdine is shown in A (1 to 100 µM) and B (1 to 10 µM). Currents were elicited by depolarizing voltage steps from –60 mV to 40 mV (the holding potential was –60 mV). Reduced currents were normalized (I/Imax) with peak currents at 40 mV before drug application. Normalized currents–voltage curves are shown in Ae and Bd. Marks above the current traces indicate the measuring points and linopirdine concentrations.
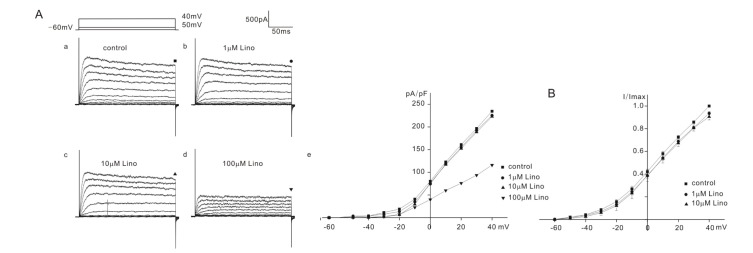
Fig. 4 Effect of linopirdine on K+ currents in ICR mice OHCs.The varying degree of inhibition of K+ currents by linopirdine is shown in A (1 to 100 µM) and B (1 to 10 µM). Currents were elicited by depolarizing voltage steps from –60 mV to 40 mV (the holding potential was –60 mV). Reduced currents were normalized (I/Imax) with peak currents at 40 mV before drug application. Normalized currents-voltage curves are shown in Ae and B. Marks above the current traces indicate the measuring points and linopirdine concentrations. Fig. 4Aa~4Ae show the linopirdine effects recorded in one cell. Fig. 4B shows normalized currents-voltage curves obtained from the cells treated with linopirdine up to 10 µM.
Reference
-
1. Marcotti W, Johnson SL, Holley MC, Kros CJ. Developmental changes in the expression of potassium currents of embryonic, neonatal and mature mouse inner hair cells. J Physiol. 2003; 548:383–400. PMID: 12588897.
Article2. Marcotti W, Géléoc GS, Lennan GW, Kros CJ. Transient expression of an inwardly rectifying potassium conductance in developing inner and outer hair cells along the mouse cochlea. Pflugers Arch. 1999; 439:113–122. PMID: 10651007.
Article3. Dulon D, Luo L, Zhang C, Ryan AF. Expression of small-conductance calcium-activated potassium channels (SK) in outer hair cells of the rat cochlea. Eur J Neurosci. 1998; 10:907–915. PMID: 9753158.
Article4. Roux I, Wersinger E, McIntosh JM, Fuchs PA, Glowatzki E. Onset of cholinergic efferent synaptic function in sensory hair cells of the rat cochlea. J Neurosci. 2011; 31:15092–15101. PMID: 22016543.
Article5. Marcotti W, Kros CJ. Developmental expression of the potassium current IK,n contributes to maturation of mouse outer hair cells. J Physiol. 1999; 520:653–660. PMID: 10545133.6. Kros CJ, Ruppersberg JP, Rüsch A. Expression of a potassium current in inner hair cells during development of hearing in mice. Nature. 1998; 394:281–284. PMID: 9685158.
Article7. Ahn JW, Kang SW, Ahn SC. Characteristics of K+ outward currents in the cochlear outer hair cells of circling mice within the first postnatal week. Korean J Physiol Pharmacol. 2015; 19:383–388. PMID: 26170743.8. Chung WH, Kim KR, Cho YS, Cho DY, Woo JH, Ryoo ZY, Cho KI, Hong SH. Cochlear pathology of the circling mouse: a new mouse model of DFNB6. Acta Otolaryngol. 2007; 127:244–251. PMID: 17364360.
Article9. Lee JW, Lee EJ, Hong SH, Chung WH, Lee HT, Lee TW, Lee JR, Kim HT, Suh JG, Kim TY, Ryoo ZY. Circling mouse: possible animal model for deafness. Comp Med. 2001; 51:550–554. PMID: 11924819.10. Lee Y, Chang SY, Jung JY, Ahn SC. Reinvestigation of cochlear pathology in circling mice. Neurosci Lett. 2015; 594:30–35. PMID: 25817368.
Article11. Hong SH, Kim MJ, Ahn SC. Glutamatergic transmission is sustained at a later period of development of medial nucleus of the trapezoid body-lateral superior olive synapses in circling mice. J Neurosci. 2008; 28:13003–13007. PMID: 19036993.
Article12. Helyer RJ, Kennedy HJ, Davies D, Holley MC, Kros CJ. Development of outward potassium currents in inner and outer hair cells from the embryonic mouse cochlea. Audiol Neurootol. 2005; 10:22–34. PMID: 15486441.
Article13. Pattnaik BR, Hughes BA. Effects of KCNQ channel modulators on the M-type potassium current in primate retinal pigment epithelium. Am J Physiol Cell Physiol. 2012; 302:C821–C833. PMID: 22135213.
Article14. Søgaard R, Ljungstrøm T, Pedersen KA, Olesen SP, Jensen BS. KCNQ4 channels expressed in mammalian cells: functional characteristics and pharmacology. Am J Physiol Cell Physiol. 2001; 280:C859–C866. PMID: 11245603.
Article15. Wang HS, Pan Z, Shi W, Brown BS, Wymore RS, Cohen IS, Dixon JE, McKinnon D. KCNQ2 and KCNQ3 potassium channel subunits: molecular correlates of the M-channel. Science. 1998; 282:1890–1893. PMID: 9836639.
Article16. Schwarz JR, Glassmeier G, Cooper EC, Kao TC, Nodera H, Tabuena D, Kaji R, Bostock H. KCNQ channels mediate IKs, a slow K+ current regulating excitability in the rat node of Ranvier. J Physiol. 2006; 573:17–34. PMID: 16527853.17. Housley GD, Ashmore JF. Ionic currents of outer hair cells isolated from the guinea-pig cochlea. J Physiol. 1992; 448:73–98. PMID: 1593487.
Article18. Kimitsuki T, Komune N, Noda T, Takaiwa K, Ohashi M, Komune S. Property of IK,n in inner hair cells isolated from guinea-pig cochlea. Hear Res. 2010; 261:57–62. PMID: 20060884.19. Bal M, Zhang J, Zaika O, Hernandez CC, Shapiro MS. Homomeric and heteromeric assembly of KCNQ (Kv7) K+ channels assayed by total internal reflection fluorescence/fluorescence resonance energy transfer and patch clamp analysis. J Biol Chem. 2008; 283:30668–30676. PMID: 18786918.20. Robbins J. KCNQ potassium channels: physiology, pathophysiology, and pharmacology. Pharmacol Ther. 2001; 90:1–19. PMID: 11448722.
Article21. Schnee ME, Brown BS. Selectivity of linopirdine (DuP 996), a neurotransmitter release enhancer, in blocking voltage-dependent and calcium-activated potassium currents in hippocampal neurons. J Pharmacol Exp Ther. 1998; 286:709–717. PMID: 9694925.22. Lamas JA, Selyanko AA, Brown DA. Effects of a cognition-enhancer, linopirdine (DuP 996), on M-type potassium currents (IK(M)) and some other voltage- and ligand-gated membrane currents in rat sympathetic neurons. Eur J Neurosci. 1997; 9:605–616. PMID: 9104602.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Characteristics of K+ Outward Currents in the Cochlear Outer Hair Cells of Circling Mice within the First Postnatal Week
- Over-expression of myosin7A in cochlear hair cells of circling mice
- Temperature Enhances Activation and Inactivation Kinetics of Potassium Currents in Inner Hair Cells Isolated from Guinea-Pig Cochlea
- Mechanism of Efferent Inhibition in Cochlear Hair Cell
- Changes of Cochlear Nerve Terminals after Temporary Noise-Induced Hearing Loss
