Sox9 Increases the Proliferation and Colony-forming Activity of Outer Root Sheath Cells Cultured In Vitro
- Affiliations
-
- 1Department of Dermatology and Research Institute for Medical Sciences, School of Medicine, Chungnam National University, Daejeon, Korea. cdkimd@cnu.ac.kr
- 2Department of Nursing, Kongju National University, Gongju, Korea.
- KMID: 2156653
- DOI: http://doi.org/10.5021/ad.2011.23.2.138
Abstract
- BACKGROUND
beta-catenin plays a pivotal role in hair follicle development and hair growth cycle.
OBJECTIVE
The aim of this study was to identify beta-catenin-regulated genes in cultured human hair outer root sheath (ORS) cells.
METHODS
Primary cultured ORS cells were transduced with recombinant adenovirus expressing N-terminal truncated beta-catenin (constitutive active form), and beta-catenin-regulated genes were identified.
RESULTS
Overexpression of the constitutively active form of beta-catenin led to induction of Sox9 expression at both mRNA and protein levels. To investigate the potential role of Sox9, we made the recombinant adenovirus expressing green fluorescent protein-tagged Sox9, and then transduced into cultured ORS cells. Interestingly, Sox9 induced the expression of keratin 15, increased the proliferation of ORS cells in vitro, and enhanced colony-forming activity.
CONCLUSION
Our results suggest that Sox9 is a beta-catenin-regulated gene in ORS cells, and has potential importance in the regulation of hair follicle homeostasis.
Keyword
MeSH Terms
Figure
-
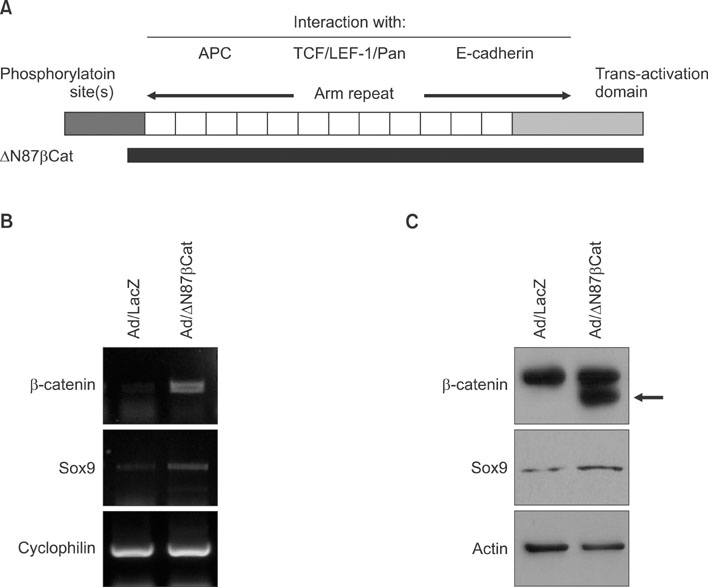
Fig. 1 (A) Structure of β-catenin. The N-terminus has several phosphorylation sites, which are important for regulating the stability of β-catenin. Central domain consists of 12 arm repeats, which allow for overlapping interaction with multiple binding partners. The C-terminus functions as a transcriptional activation domain. N-terminal 87-amino acid truncated β-catenin (ΔN87β Cat) is depicted under the wild-type β-catenin. (B) Outer root sheath cells were transduced with adenovirus expressing ΔN87βCat at 10 multiplicity of infection. The mRNA level was verified by reverse transcription-polymerase chain reaction. (C) After adenoviral transduction, protein level was verified by Western blot. Upper band represents the endogenous β-catenin, while lower band represents the N-truncated β-catenin (arrow). Adenovirus expressing lacZ was used as negative control.

Fig. 2 Immunohistochemical staining of Sox9 (×100). Paraffin section of scalp specimen was incubated with anti-Sox9 antibody, and then sequentially incubated with HRP-conjugated secondary antibody. Intense expression of Sox9 is detected in the outer layer of outer root sheath in middle portion of hair follicle.

Fig. 3 (A) Expression of exogenous Sox9 in outer root sheath (ORS) cells. ORS cells were transduced with adenovirus expressing GFP-tagged Sox9 (GFP-Sox9) at 10 multiplicity of infection (MOI). Adenovirus expressing GFP was used as negative control. Twenty-four hours after adenoviral transduction, cells were observed under the fluorescent microscopy. (B) Effect of Sox9 on the expression of stem cell marker in ORS cells. Cells were transduced with adenovirus expressing GFP-Sox9 at the indicated MOI. The mRNA level was verified by reverse transcription-polymerase chain reaction. (C) Cellular extracts were prepared, and the protein level for stem cell marker was verified by Western blot. Actin was used as a loading control.
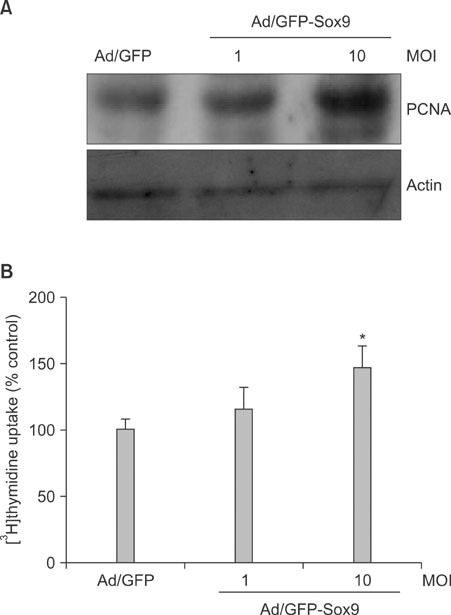
Fig. 4 Effect of Sox9 on cell proliferation. (A) Cells were transduced with adenovirus expressing GFP-Sox9 at the indicated multiplicity of infection. The proliferation marker PCNA was examined by Western blot. (B) Thymidine uptake assay. After adenoviral transduction, cells were received [3H]thymidine-containing medium. After a 3 day incubation, cells were lysed and radioactivity was measured using liquid scintillation counter. Data are expressed as % control and SEM from three independent experiments. *p<0.01.
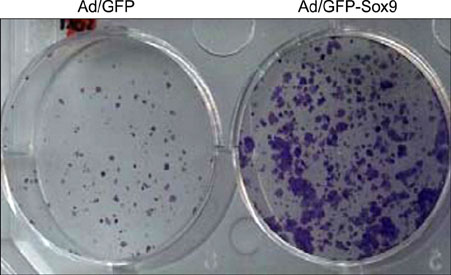
Fig. 5 Effect of Sox9 on clonogenicity. ORS cells were transduced with adenovirus (10 multiplicity of infection). After 2 day incubation, cells were detached from the culture dish and 1,000 cells were re-seeded onto new culture dish. After 2 week incubation, colony formation was verified by staining with Crystal violet.
Reference
-
1. Paus R, Cotsarelis G. The biology of hair follicles. N Engl J Med. 1999. 341:491–497.
Article2. Hülsken J, Behrens J. The Wnt signalling pathway. J Cell Sci. 2000. 113:3545.
Article3. Gat U, DasGupta R, Degenstein L, Fuchs E. De Novo hair follicle morphogenesis and hair tumors in mice expressing a truncated β-catenin in skin. Cell. 1998. 95:605–614.
Article4. Huelsken J, Vogel R, Erdmann B, Cotsarelis G, Birchmeier W. β-Catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell. 2001. 105:533–545.
Article5. Sohn KC, Shi G, Jang S, Choi DK, Lee Y, Yoon TJ, et al. Pitx2, a β-catenin-regulated transcription factor, regulates the differentiation of outer root sheath cells cultured in vitro. J Dermatol Sci. 2009. 54:6–11.
Article6. Kim JH, Choi DK, Lee SS, Choi SJ, Kim CD, Yoon TJ, et al. Enhancement of keratinocyte differentiation by rose absolute oil. Ann Dermatol. 2010. 22:255–261.
Article7. Wong MH, Rubinfeld B, Gordon JI. Effects of forced expression of an NH2-terminal truncated β-catenin on mouse intestinal epithelial homeostasis. J Cell Biol. 1998. 141:765–777.
Article8. Vidal VP, Chaboissier MC, Lützkendorf S, Cotsarelis G, Mill P, Hui CC, et al. Sox9 is essential for outer root sheath differentiation and the formation of the hair stem cell compartment. Curr Biol. 2005. 15:1340–1351.
Article9. Ohyama M, Terunuma A, Tock CL, Radonovich MF, Pise-Masison CA, Hopping SB, et al. Characterization and isolation of stem cell-enriched human hair follicle bulge cells. J Clin Invest. 2006. 116:249–260.
Article10. Yang JS, Lavker RM, Sun TT. Upper human hair follicle contains a subpopulation of keratinocytes with superior in vitro proliferative potential. J Invest Dermatol. 1993. 101:652–659.
Article11. Claudinot S, Nicolas M, Oshima H, Rochat A, Barrandon Y. Long-term renewal of hair follicles from clonogenic multipotent stem cells. Proc Natl Acad Sci U S A. 2005. 102:14677–14682.
Article12. Cotsarelis G, Sun TT, Lavker RM. Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell. 1990. 61:1329–1337.
Article13. Van Mater D, Kolligs FT, Dlugosz AA, Fearon ER. Transient activation of β-catenin signaling in cutaneous keratinocytes is sufficient to trigger the active growth phase of the hair cycle in mice. Genes Dev. 2003. 17:1219–1224.
Article14. Barrionuevo F, Scherer G. SOX E genes: SOX9 and SOX8 in mammalian testis development. Int J Biochem Cell Biol. 2010. 42:433–436.
Article15. Yano F, Kugimiya F, Ohba S, Ikeda T, Chikuda H, Ogasawara T, et al. The canonical Wnt signaling pathway promotes chondrocyte differentiation in a Sox9-dependent manner. Biochem Biophys Res Commun. 2005. 333:1300–1308.
Article16. Blache P, van de, Duluc I, Domon C, Berta P, Freund JN, et al. SOX9 is an intestine crypt transcription factor, is regulated by the Wnt pathway, and represses the CDX2 and MUC2 genes. J Cell Biol. 2004. 166:37–47.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Growth Effect of Minoxidil and Minoxidil Sulfate on Cultured Human Keratinocytes and Outer Root Sheath Cells
- Expression of p63 and its association with cell proliferation at different stages of murine hair follicle cycle
- Dieckol Inhibits the Effects of Particulate Matter 10 on Sebocytes, Outer Root Sheath Cells, and Cutibacterium Acnes−Pretreated Mice
- Effects of Black Ginseng Water Extract under the Inflammatory Conditions of Cultured Sebocytes and Outer Root Sheath Cells
- Effects of <10-μm Particulate Matter on Cultured Human Sebocytes and Outer Root Sheath Cells and Usefulness of Siegesbeckia Herba Extract
