Expression of transient receptor potential channels in the ependymal cells of the developing rat brain
- Affiliations
-
- 1Department of Neurology, Gangneung Asan Hospital, University of Ulsan College of Medicine, Gangneung, Korea.
- 2Department of Anatomy, Kwandong University College of Medicine, Gangneung, Korea. dextrin@kd.ac.kr
- 3Department of Anatomy, Yonsei University College of Medicine, Seoul, Korea.
- KMID: 2046759
- DOI: http://doi.org/10.5115/acb.2013.46.1.68
Abstract
- Cerebrospinal fluid (CSF) plays an important role in providing brain tissue with a stable internal environment as well as in absorbing mechanical and thermal stresses. From its initial composition, derived from the amniotic fluid trapped by the closure of neuropores, CSF is modified by developing and differentiating ependymal cells lining the ventricular surface or forming the choroid plexus. Its osmolarity and ionic composition brings about a change through the action of many channels expressed on the ependymal cells. Some newly discovered transient receptor potential (TRP) channels are known to be expressed in the choroid plexus ependyma. To detect additional TRP channel expression, immunohistochemical screening was performed at the choroid plexus of 13-, 15-, 17-, and 19-day embryos, using antibodies against TRPV1, TRPV3, and TRPA1, and the expression was compared with those in the adult TRP channels. The level of TRP channel expression was higher in the choroid plexus which suggests more active functioning of TRP channels in the developing choroid plexus than the ventricular lining ependyma in the 15- and 17-day embryos. All the expression of TRP channels decreased at the 19th day of gestation. TRPA1 was expressed at a higher level than TRPV1 and TRPV3 in almost all stages in both the choroid plexus and ventricular lining epithelium. The highest level of TRPV1 and TRPV3 expression was observed in association with the glycogen deposits in the cytoplasm of the choroid plexus ependymal cells of the 15- and 17-day embryos.
MeSH Terms
Figure
-
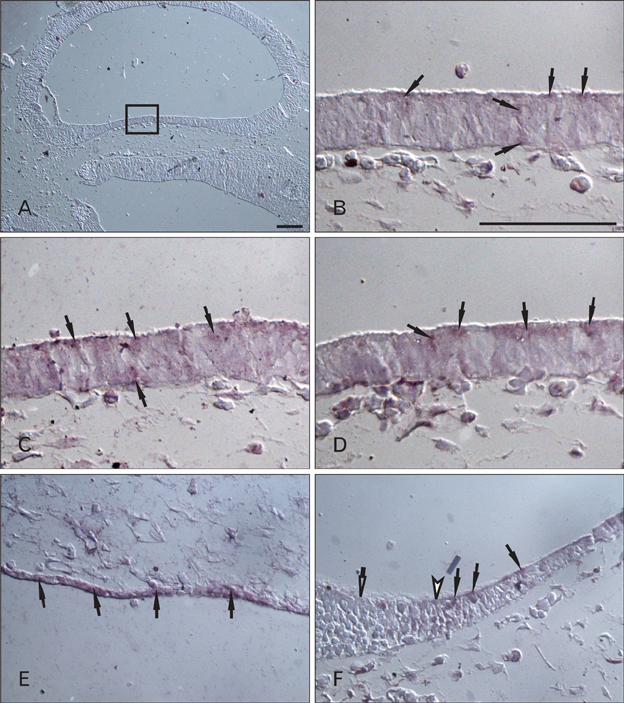
Fig. 1 Telencephalic and rhombencephalic vesicles of the 13-day rat embryo. (A) Horizontal section of the developing telencephalon just above the level of the interventricular foramen. In the magnified view of the area (B) demarcated within a rectangle in (A), weakly TRPV1-positive neuroepithelial cells are noted (arrows). Identical regions with same magnification with B, show the weakly TRPV3-positive (C, 1:200, arrows), and TRPA1-positive cells (D, 1:800, arrows), respectively. (E) TRPV3-positive (1:400, arrows) neuroepithelial cells formed a single-layered posterior wall of the rhombencephalic vesicle. These cells also showed the immunopositivity against TRPV1 and TRPA1. (F) The transitional region from the posterior wall on the right to the anterior wall on the left in the rhombencephalon. The sulcus limitans (open arrowhead) was not well demarcated. TRPA1-positive cells (1:800, filled arrows) formed the posterior wall of rhombencephalon while neuroepithelial cells lining the anterior wall did not show immunoreactivity against TRPA1 (open arrow). Scale bar in (A)=100 µm; in (B)=100 µm (B-F).
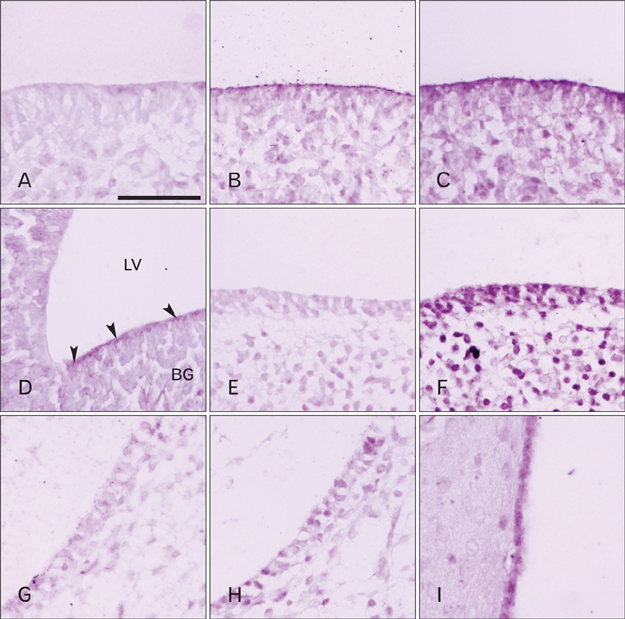
Fig. 2 Ventricular lining ependyma (VLE) from the 15-day rat embryo to the adult. In VLE of the fourth ventricle of the 15-day embryo, a diverse pattern of TRPV1 (A), TRPV3 (B), and TRPA1 (C, D) expression was noted. The expression of TRPA1 was strongest, followed by TRPV3 and that of TRPV1 was weakest. In this stage, the immunopositivity was generally observed in the apical cytoplasm. A regional difference in the expression pattern of TRPA1 in the VLE lining the developing lateral ventricles (LV) was shown in (D). Moderately TRPA1-positive VLE cells (arrowheads) covered the growing basal ganglia (BG). In the VLE of the 17-day embryo, both TRPV1-(E) and TRPV3-immunoreactivity were weak, while TRPA1-reactivity was moderate (F). The expression of all 3 TRP channels decreased in the 19-day embryo showing a weak reactivity against TRPV3 (G) and TRPA1 (H). In the adult, VLE showed a moderate level of expression of TRPV3 and TRPA1 (I). Scale bar in (A)=50 µm (A-I).

Fig. 3 Choroid plexus ependyma (CPE) in the 15- and 17-day rat embryos. Immunohistochemical staining result using antibodies against TRPV1 (A, D), TRPV3 (B, E), and TRPA1 (C, F) in the 15-day (A-C) and the 17-day (D-F) embryos. In the stroma of the 15-day series, blood vessels were not fully developed, while those of the 17-day showed large vascular lumina filled with red blood corpuscles. In the 15-day embryo glycogen droplets with oval outline began to develop accompanied with strong immunoreactivity against TRPV1 (A, arrows) and TRPV3 (B, arrows) around their periphery. These glycogen inclusions enlarged in the next series, the 17-day embryo, but lost their expression of TRPV1 (D, open arrowheads). They retained the expression of TRPV3 (E, arrows) but the immunoreactivity was found not only at the periphery of glycogen but along the basal membrane of CPE. Glycogen in some CPE show the express TRPV3. TRPA1 was not expressed in relation with the glycogen accumulations but moderate immunopositivity was observed in the apical cytoplasm of some CPE in the 15-day embryo (C, arrowheads), and in the entire cytoplasm in 17-day embryo (F, arrowheads). TRPV1 and TRPV3 were also expressed of some CPE in these stages. Scale bar in (A)=50 µm (A-F).
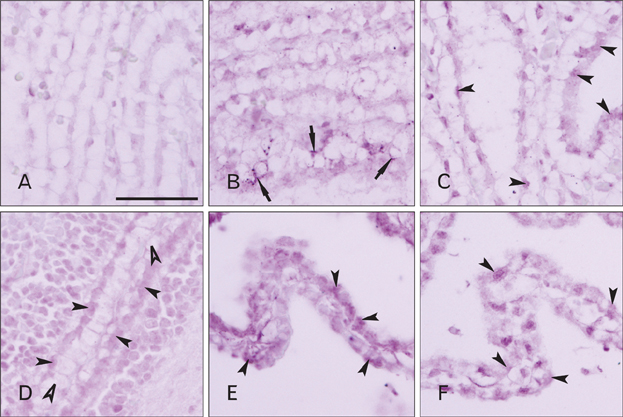
Fig. 4 Choroid plexus ependyma (CPE) in the 19-day rat embryo and in the adult. In the cytoplasm of CPE of the 19-day embryo, TRPV1 was very weakly expressed (A), while TRPV3 (B) and TRPA1 (C, arrowheads) were expressed at a higher level. Due to a dramatic shrinkage of the lateral and the third ventricles, CPE (D, arrowheads) and VLE were in close proximity in the 19-day embryo. Glycogen (D, open arrowheads) occupied a great portion of cytoplasm and a few of the droplets retained the TRPV3 expression around their peripheral cytoplasm (B, arrows). In the adult, there was a moderate degree of TRPV3 and TRPA1 expression in the cytoplasm of CPE (E and F, arrowheads), with a difference in their expression pattern. TRPV3 immunopositivity was observed in most of the CPE having some regions of strong reactivity inside their cytoplasm (E), while some CPE having moderate reactivity to TRPA1 could be recognized. Scale bar in (A)=50 µm (A-F).
Reference
-
1. Oldendorf WH, Davson H. Brain extracellular space and the sink action of cerebrospinal fluid. Measurement of rabbit brain extracellular space using sucrose labeled with carbon 14. Arch Neurol. 1967. 17:196–205.2. Di Terlizzi R, Platt S. The function, composition and analysis of cerebrospinal fluid in companion animals: part I - function and composition. Vet J. 2006. 172:422–431.3. Damkier HH, Brown PD, Praetorius J. Epithelial pathways in choroid plexus electrolyte transport. Physiology (Bethesda). 2010. 25:239–249.4. Millar ID, Bruce J, Brown PD. Ion channel diversity, channel expression and function in the choroid plexuses. Cerebrospinal Fluid Res. 2007. 4:8.5. Talavera K, Nilius B, Voets T. Neuronal TRP channels: thermometers, pathfinders and life-savers. Trends Neurosci. 2008. 31:287–295.6. Liedtke W, Choe Y, Martí-Renom MA, Bell AM, Denis CS, Sali A, Hudspeth AJ, Friedman JM, Heller S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell. 2000. 103:525–535.7. Harteneck C, Reiter B. TRP channels activated by extracellular hypo-osmoticity in epithelia. Biochem Soc Trans. 2007. 35(Pt 1):91–95.8. Benham CD, Davis JB, Randall AD. Vanilloid and TRP channels: a family of lipid-gated cation channels. Neuropharmacology. 2002. 42:873–888.9. Pedersen SF, Owsianik G, Nilius B. TRP channels: an overview. Cell Calcium. 2005. 38:233–252.10. Voets T, Talavera K, Owsianik G, Nilius B. Sensing with TRP channels. Nat Chem Biol. 2005. 1:85–92.11. Venkatachalam K, Montell C. TRP channels. Annu Rev Biochem. 2007. 76:387–417.12. Phelps CB, Wang RR, Choo SS, Gaudet R. Differential regulation of TRPV1, TRPV3, and TRPV4 sensitivity through a conserved binding site on the ankyrin repeat domain. J Biol Chem. 2010. 285:731–740.13. Vriens J, Appendino G, Nilius B. Pharmacology of vanilloid transient receptor potential cation channels. Mol Pharmacol. 2009. 75:1262–1279.14. Das GD. Gliogenesis and ependymogenesis during embryonic development of the rat. An autoradiographic study. J Neurol Sci. 1979. 43:193–204.15. Kilinc M, Deniz M, Ketani S, Hatipoglu ES. Prenatal and postnatal development of the rat choroid plexus. J Anim Vet Adv. 2010. 9:12–15.16. Bayer SA, Altman J, Russo RJ, Zhang X. Timetables of neurogenesis in the human brain based on experimentally deter mined patterns in the rat. Neurotoxicology. 1993. 14:83–144.17. Tennyson VM, Pappas GD. Fine structure of the developing telencephalic and myelencephalic choroid plexus in the rabbit. J Comp Neurol. 1964. 123:379–411.18. Dziegielewska KM, Ek J, Habgood MD, Saunders NR. Development of the choroid plexus. Microsc Res Tech. 2001. 52:5–20.19. Johansson PA, Dziegielewska KM, Ek CJ, Habgood MD, Møllgård K, Potter A, Schuliga M, Saunders NR. Aquaporin-1 in the choroid plexuses of developing mammalian brain. Cell Tissue Res. 2005. 322:353–364.20. de Rougemont , Ames A 3rd, Nesbett FB, Hofmann HF. Fluid formed by choroid plexus; a technique for its collection and a comparison of its electrolyte composition with serum and cisternal fluids. J Neurophysiol. 1960. 23:485–495.21. Oresković D, Klarica M. The formation of cerebrospinal fluid: nearly a hundred years of interpretations and misinterpretations. Brain Res Rev. 2010. 64:241–262.22. Liddelow SA, Temple S, Møllgård K, Gehwolf R, Wagner A, Bauer H, Bauer HC, Phoenix TN, Dziegielewska KM, Saunders NR. Molecular characterisation of transport mechanisms at the developing mouse blood-CSF interface: a transcriptome approach. PLoS One. 2012. 7:e33554.23. Goldstein MH, Bazer FW, Barron DH. Characterization of changes in volume, osmolarity and electrolyte composition of porcine fetal fluids during gestation. Biol Reprod. 1980. 22:1168–1180.24. Davson H, Purvis C. Cryoscopic apparatus suitable for studies on aqueous humour and cerebro-spinal fluid. J Physiol. 1954. 124:12P–13P.25. Ames A 3rd, Sakanoue M, Endo S. Na, K, Ca, Mg, and C1 concentrations in choroid plexus fluid and cisternal fluid compared with plasma ultrafiltrate. J Neurophysiol. 1964. 27:672–681.26. Hughes AL, Pakhomova A, Brown PD. Regulatory volume increase in epithelial cells isolated from the mouse fourth ventricle choroid plexus involves Na(+)-H(+) exchange but not Na(+)-K(+)-2Cl(-) cotransport. Brain Res. 2010. 1323:1–10.27. Nilius B, Owsianik G, Voets T, Peters JA. Transient receptor potential cation channels in disease. Physiol Rev. 2007. 87:165–217.28. Bayer SA, Altman J, Russo RJ, Dai XF, Simmons JA. Cell migration in the rat embryonic neocortex. J Comp Neurol. 1991. 307:499–516.29. Desmond ME, Jacobson AG. Embryonic brain enlargement requires cerebrospinal fluid pressure. Dev Biol. 1977. 57:188–198.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Role of Transient Receptor Potential Channel in Pain
- Immunohistochemical Study on the Distribution of Canonical Transient Receptor Potential Channels in Rat Cerebellum
- Immunohistochemical Changes of Nestin, BrdU, and GFAP in the Cells of Rat Spinal Ependymal Zone According to Their Developmental Stages
- Identification of phospholipase C β downstream effect on transient receptor potential canonical 1/4, transient receptor potential canonical 1/5 channels
- Modulation of Dopaminergic Neuronal Excitability by Zinc through the Regulation of Calcium-related Channels
