Effects of pH on Vascular Tone in Rabbit Basilar Arteries
- Affiliations
-
- 1Department of Physiology, Chungbuk National University College of Medicine, Cheongju, Korea. kimkw@plaza.snu.ac.kr
- 2Department of Physiology and Biophysics, Seoul National University College of Medicine, Seoul, Korea.
- KMID: 1785692
- DOI: http://doi.org/10.3346/jkms.2004.19.1.42
Abstract
- Effects of pH on vascular tone and L-type Ca2+ channels were investigated using Mulvany myograph and voltage-clamp technique in rabbit basilar arteries. In rabbitbasilar arteries, high K+ produced tonic contractions by 11+/-0.6 mN (mean+/-S.E., n=19). When extracellular pH (pHo) was changed from control 7.4 to 7.9 ([alkalosis]o), K+-induced contraction was increased to 128+/-2.1% of the control (n=13). However, K+-induced contraction was decreased to 73+/-1.3% of the control at pHo 6.8 ([acidosis]o, n=4). Histamine (10 micrometer) also produced tonic contraction by 11+/-0.6 mN (n=17), which was blocked by post-application of nicardipine (1 micrometer). [alkalosis]o and [acidosis]o increased or decreased histamine-induced contraction to 134+/-5.7% and 27+/-7.6% of the control (n=4, 6). Since high K+- and histamine-induced tonic contractions were affected by nicardipine and pHo, the effect of pHo on voltage-dependent L-type Ca2+ channel (VDCCL) was studied. VDCCL was modulated by pHo: the peak value of Ca2+ channel current (IBa) at a holding of 0 mV decreased in [acidosis]o by 41+/-8.8%, whereas that increased in [alkalosis]o by 35+/-2.1% (n=3). These results suggested that the external pH regulates vascular tone partly via the modulation of VDCC in rabbit basilar arteries.
Keyword
MeSH Terms
-
Animals
Arteries/*pathology
Basilar Artery/*pathology
Calcium/metabolism
Calcium Channels/chemistry
Electrophysiology
Histamine/chemistry/metabolism
Hydrogen-Ion Concentration
Muscle Cells/cytology
Muscle Contraction
Muscle, Smooth/*pathology
Patch-Clamp Techniques
Potassium/chemistry/metabolism
Rabbits
Stress, Mechanical
Time Factors
Figure
-

Fig. 1 Effects of the alteration of the extracellular pH (pHo) on high K+-induced contraction in basilar arteries of rabbits. Mulvany myograph was used for recording isometric tension of rabbit basilar artery. All contractile experiments in this study were done in the presence of nitro-L-arginine (L-NNA, 100 µM). (A) Superfusion of 50 mM of high K+ solution to the bath provoked tonic contraction. This contraction was enhanced by pHo 7.9 in a reversible manner. (B, C) Under acidic condition such as pHo 7.0 or 6.8 tonic contraction was suppressed in a reversible manner. (D) Bar graphs show mean relative K+-induced contraction by alteration of pHo. Asterisks indicate the data which were considered to be significantly different from control data (**p<0.01).

Fig. 2 Effects of the repetitive alteration of pHo on high K+-induced contraction in rabbit basilar artery. Modulatory effects on high K+-induced contraction by the alteration of pHo was monitored by repetitive changes of pHo in rabbit basilar artery. (A) 50 mM of high K+-induced tonic contraction was repeatedly increased by pHo 7.9. (B) Acidotic and alkalotic conditions were induced to 50 mM of high K+-induced tonic contraction. Each condition of pHo 6.8 and 7.9 in a same tissue decreased and increased tonic contraction, respectively, in a reversible manner. (C) In the presence of L-NNA, enhancing effects of alkalotic condition on high K+-induced tonic contraction was studied by pre-application of pHo 7.9. Tonic contraction induced by application of 50 mM of high K+ solution (pHo 7.9) was decreased by post application of normal pH (pH 7.4).

Fig. 3 Histamine-induced contraction in rabbit basilar artery. (A) Histamine (1-20 µM) produced contractions in a concentration-dependent manner. Approximately, 10 µM of histamine produced maximal contraction. (B) In some cases, application of histamine evoked regular small oscillatory phasic contractions. (C) Histamine-induced tonic contraction was suppressed by post-application of nicardipine (1 µM).

Fig. 4 Effects of alteration of pHo on the histamine-induced contraction in basilar arteries of rabbits. 10 µM of histamine produced tonic contractions in rabbit basilar artery. (A, B) Histamine-induced contraction was decreased or increased by pHo 6.8 or pHo 7.9 in a reversible manner. (C) Bar graphs show mean relative histamine-induced contraction by alteration of pHo. Asterisks indicate the data which were considered to be significantly different from control data (**p<0.01).
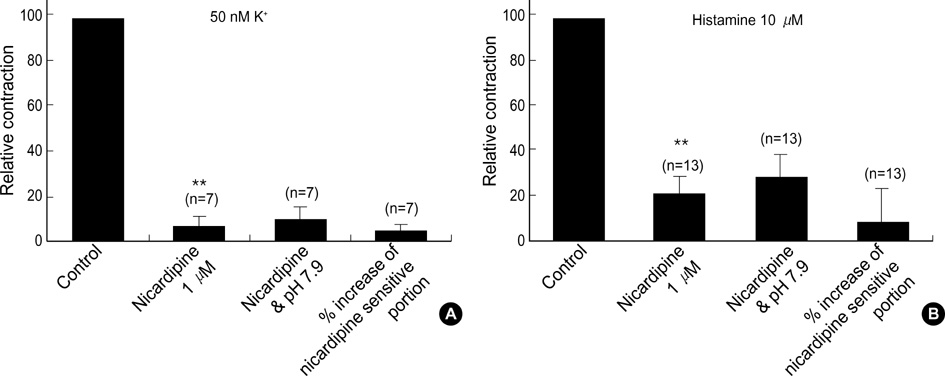
Fig. 5 Effects of pHo in the presence of nicardipine on High K+- and histamine-induced Contraction in Rabbit Basilar Arteries. In A, B, effects of nicardipine and [alkalosis]o in the presence of nicardipine on high K+- and histamine-induced contractions were summarized. Pannel A shows nicardipine completely suppressed high K+-induced sustained contraction and these effect was not reversed by [alkalosis]o significantly. In pannel B, nicardipine suppressed histamine-induced sustained contraction and it was not reversed by [alkalosis]o significantly. Asterisks indicate the data which were considered to be significantly different from control data (**p<0.01).
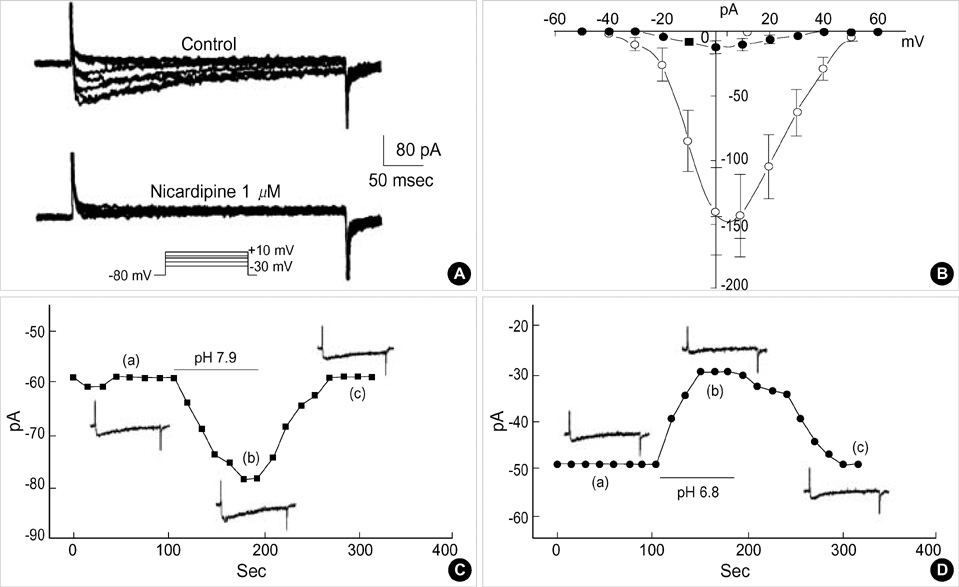
Fig. 6 Effects of alteration of pHo on Ca2+ channel current (IBa) in rabbit basilar artery. The membrane potential was held at -80 mV and 2 mM Ca2+ was used as a charge carrier. Step 10 mV depolarizing pulses from -50 to +60 mV were applied for 500 msec. (A) Depolarizing pulses above -30 mV elicited inward currents (ICa) and nicardipine (1 µM) almost completely blocked ICa. (B) Current/voltage (I/V) relationships of ICa were obtained in the absence and presence of nicardipine (1 µM). In I/V relationships, ICa was significantly suppressed by nicardipine throughout the whole test potential range. 10 mM Ba2+ was used as a charge carrier for the study of regulation of Ca2+ channel current (IBa) by pHo. In C, D effects of alteration of pHo on IBa were studied. Step depolarizing pulse from -80 to 0 mV were applied for 500 msec every 15 sec. When normal external solution (pHo 7.4) was changed to pHo 7.9, IBa was increased in a reversible manner (raw traces of IBa, see insets). However, pHo 6.8 decreased IBa in a reversible manner (raw traces of IBa, insets).
Reference
-
1. Tian R, Vogel P, Lassen NA, Mulvany MJ, Andreasen F, Aalkjaer C. Role of extracellular and intracellular acidosis for hypercapnia-induced inhibition of tension of isolated rat cerebral arteries. Circ Res. 1995. 76:269–275.
Article2. Kontos HA. Regulation of the cerebral circulation. Annu Rev Physiol. 1981. 43:397–407.
Article3. Toda N, Hatano Y, Mori K. Mechanisms underlying response to hypercapnia and bicarbonate of isolated dog cerebral arteries. Am J Physiol. 1989. 257:H141–H146.
Article4. Klöckner U, Isenberg G. Intracellular pH modulates the avalability of vascular L-type Ca2+ channels. J Gen Physiol. 1994. 103:647–663.5. Klöckner U, Isenberg G. Calcium channel current of vascular smooth muscle cells: extracellular protons modulate gating and single channel conductance. J Gen Physiol. 1994. 103:665–678.
Article6. Smith JB, Dwyer SD, Smith L. Lowering extracellular pH evokes inositol polyphosphate formation and calcium mobilization. J Biol Chem. 1989. 264:8723–8728.
Article7. Ishizaka H, Kuo L. Acidosis-induced coronary arteriolar dilation is mediated by ATP-sensitive potassium channels in vascular smooth muscle. Circ Res. 1996. 78:50–57.
Article8. Gokina NI, Bevan JA. Histamine-induced depolarization: ionic mechanisms and role in sustained contraction of rabbit cerebral arteries. Am J Physiol Heart Circ Physiol. 2000. 278:H2094–H2104.9. Gokina NI, Bevan JA. Role of intracellular Ca2+ release in histamine-induced depolarization in rabbit middle cerebral artery. Am J Physiol Heart Circ Physiol. 2000. 278:H2105–H2114.10. Austin C, Wray S. The effects of extracellular pH and calcium change on force and intracellular calcium in rat vascular smooth muscle. J Physiol. 1995. 488:281–291.
Article11. Rinaldi CJ, Amando Cattaneo E, Cigolani HE. Interaction between calcium and hydrogen ions in canine coronary arteries. J Mol Cell Cardiol. 1987. 19:773–784.12. Peng H-L, Jensen PE, Nilsson H, Aalkjaer C. Effect of acidosis on tension and [Ca2+]i in rat cerebral arteries: is there a role for membrane potential? Am J Physiol. 1998. 274:H655–H662.13. Oike M, Inoue Y, Kitamura K, Kuriyama H. Dual action of FRC8653, a novel dihydropyridine derivative, on the Ba2+ current recorded from the rabbit basilar artery. Circ Res. 1990. 67:993–1006.14. Tsien RW, Ellinor PT, Horne WA. Molecular diversity of voltage-dependent Ca2+ channels. Trends Pharmacol Sci. 1991. 12:349–354.15. Worley JF, Quayle JM, Standen NB, Nelson MT. Regulation of single calcium channels in cerebral arteries by voltage, serotonin, and dihydropyridines. Am J Physiol. 1991. 261:H1951–H1960.
Article16. West GA, Leppla DC, Simard JM. Effects of external pH on ionic currents in smooth muscle cells from the basilar artery of the guinea pig. Circ Res. 1992. 71:201–209.
Article17. Horie S, Yano S, Watanabe K. Intracellular alkalinization by NH4Cl increases cytosolic Ca2+ level and tension in the rat aortic smooth muscle. Life Sci. 1995. 56:1835–1843.18. Aoyama Y, Ueda K, Setogawa A, Kawai Y. Effects of pH on contraction and Ca2+ mobilization in vascular smooth muscles of the rabbit basilar artery. Jpn J Physiol. 1999. 49:55–62.19. Hamil OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981. 391:85–100.20. Horn R, Marty A. Muscarinic activation of ionic currents measured by a new whole-cell recording method. J Gen Physiol. 1988. 92:145–159.
Article21. Isenberg G, Klöckner U. Calcium tolerant ventricular myocytes delivered by pre-incubation in a "KB-medium". Pflugers Arch. 1982. 395:6–18.22. Suh SH, Han JJ, Park SJ, Choi JY, Sim JH, Kim YC, Kim KW. Differentmechanisms for K+-induced relaxation in various artery. Korean J Physiol Pharmacol. 1998. 3:415–425.23. So I, Kang TM, Kim KW. Characteristics of Ca currents in rabbit basilar arterial smooth muscle cells. Seoul J Med. 1994. 35:169–182.24. Bolton TB. Mechanisms of action of transmitters and other substances on smooth muscle. Physiol Rev. 1979. 59:606–719.
Article25. Carl A, Lee HK, Sanders KM. Regulation of ion channels in smooth muscles by calcium. Am J Physiol. 1996. 271:C9–C34.
Article26. Kuriyama H, Kitamura K, Nabata H. Pharmacological and physiological significance of ion channels and factors that modulate them in vascular tissues. Pharmacol Rev. 1995. 47:387–573.27. Nelson MT, Patlak JB, Worley JF, Standen NB. Calcium channels, potassium channels, and voltage dependence of arterial smooth muscle tone. Am J Physiol. 1990. 259:C3–C18.
Article28. Garland CJ. The role of membrane depolarization in the contractile response of rabbit basilar arery to 5-hydroxytryptamine. J Physiol (Lond). 1987. 392:333–348.29. Karashima T, Kuriyama H. Electrical properties of smooth muscle cell membrane and neuromuscular transmission in the guinea-pig basilar artery. Br J Pharmacol. 1991. 74:495–504.
Article30. Sürprenant A, Neild TO, Holman ME. Membrane properties of rabbit basilar arteries and their responses to transmural stimulation. Pflugers Arch. 1987. 410:92–101.31. Ishikawa T, Hume JR, Keef KD. Modulation of K and Ca2+ channels by histamine H1-receptor stimulation in rabbit coronary artery cells. J Physiol. 1993. 468:379–400.32. Oike M, Kitamura K, Kuriyama H. Histamine H3-receptor activation augments voltage-dependent Ca2+ current via GTP hydrolysis in rabbitsaphenous artery. J Physiol. 1992. 448:133–152.33. Takagi T, Tan EC, Shibata S. Characteristics of histamine receptors in human cerebral arteries. Neurol Med Chir (Tokyo). 1993. 33:675–681.
Article34. Hayabuchi Y, Nakaya Y, Matsuoka S, Kuroda Y. Effect of acidosis on Ca2+-activated K+ channels in cultured porcine coronary artery smooth muscle cells. Pflugers Arch. 1998. 436:509–514.35. Shimamura K, Sekiguchi F, Sunano S. Tension oscillation in arteries and its abnormality in hypertensive animals. Clin Exp Pharmacol Physiol. 1999. 26:275–284.
Article36. Tostes RC, Storm DS, Chi DH, Webb RC. Intracellular calcium stores and oscillatory contractions in arteries from genetically hypertensive rats. Hypertens Res. 1996. 19:103–111.
Article37. Kang TM, So I, Kim KW. Caffeine- and histamine-induced oscillatons of K (Ca) current in single smooth muscle cells of rabbit cerebral artery. Pflugers Arch. 1995. 431:91–100.38. Gurevicius J, Salem MR, Metwally AA, Silver JM, Crystal GJ. Contribution of nitric oxide to coronary vasodilation during hypercapnic acidosis. Am J Physiol. 1995. 268(1 Pt 2):H39–H47.
Article39. Cowan CL, Cohen RA. Two mechanisms mediate relaxation by bradykinin of pig coronary artery: NO-dependent and independent responses. Am J Physiol. 1991. 261:H830–H835.
Article40. Vanhoutte PM. The end of the quest? Nature. 1987. 327:459–460.
Article41. Vanhoutte PM. Other endothelium-derived vasoactive factors. Circulation. 1993. 87:Suppl V. V9–V17.42. Nagao T, Vanhoutte PM. Hyperpolarization as a mechanism for endothelium-dependent relaxations in the porcine coronary artery. J Physiol. 1992. 445:355–367.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- he Changes of the Basilar Artery Following Experimental Cerebral Subarachnoidal Hemorrhage
- Microsurgical Anatomy of the Basilar Artery: Surgical Approaches to the Basilar Trunk and Vertebrobasilar Junction Aneurysms
- A Case of Moyamoya Disease Associated with Complete Duplication of the Basilar artery
- Changes in Myogenic Tone in Spontaneously Hypertensive Rat: Role of RhoA and Protein Kinase C
- The Effect of Low-dose Papaverine on In-vivo Rabbit Basilar Artery
