Inhibition of the Human Ether-a-go-go-related Gene (HERG) K+ Channels by Lindera erythrocarpa
- Affiliations
-
- 1Department of Physiology, Institute of Bioscience and Biotechnology, Kangwon National University College of Medicine, Chuncheon, Korea. suhyunjo@kangwon.ac.kr
- 2Department of Pharmacology, Cheju National University College of Medicine, Jeju, Korea.
- 3College of Pharmacy, Chungnam National University, Daejeon, Korea.
- KMID: 1783138
- DOI: http://doi.org/10.3346/jkms.2009.24.6.1089
Abstract
- Lindera erythrocarpa Makino (Lauraceae) is used as a traditional medicine for analgesic, antidote, and antibacterial purposes and shows anti-tumor activity. We studied the effects of Lindera erythrocarpa on the human ether-a-go-go-related gene (HERG) channel, which appears of importance in favoring cancer progression in vivo and determining cardiac action potential duration. Application of MeOH extract of Lindera erythrocarpa showed a dose-dependent decrease in the amplitudes of the outward currents measured at the end of the pulse (I(HERG)) and the tail currents of HERG (I(tail)). When the BuOH fraction and H2O fraction of Lindera erythrocarpa were added to the perfusate, both I(HERG) and I(tail) were suppressed, while the hexane fraction, CHCl3 fraction, and EtOAc fraction did not inhibit either I(HERG) or I(tail). The potential required for half-maximal activation caused by EtOAc fraction, BuOH fraction, and H2O fraction shifted significantly. The BuOH fraction and H2O fraction (100 microgram/mL) decreased gmax by 59.6% and 52.9%, respectively. The H2O fraction- and BuOH fraction-induced blockades of I(tail) progressively decreased with increasing depolarization, showing the voltage-dependent block. Our findings suggest that Lindera erythrocarpa, a traditional medicine, blocks HERG channel, which could contribute to its anticancer and cardiac arrhythmogenic effect.
MeSH Terms
Figure
-
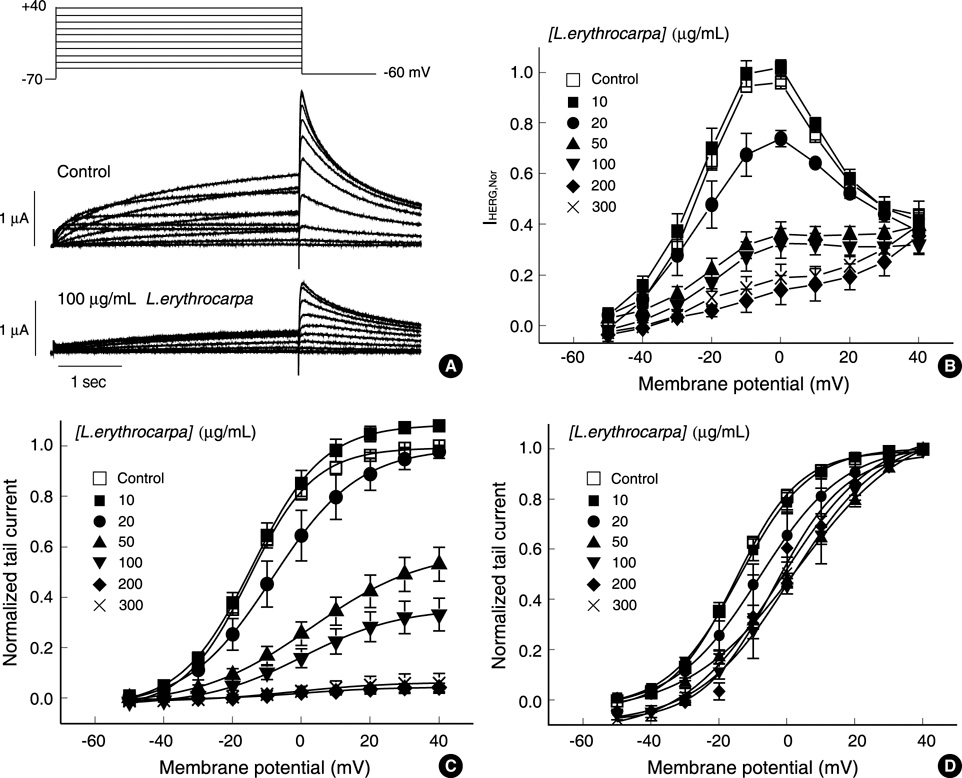
Fig. 1 Effect of Lindera erythrocarpa on human-ether-a-go-go-related-gene (HERG) currents elicited by depolarizing voltage pulses. (A) Superimposed current traces elicited by depolarizing voltage pulses (4 sec) in 10 mV steps (upper panel) from a holding potential of -70 mV in the absence of L. erythrocarpa extract (control, middle panel) and in the presence of 100 µg/mL L. erythrocarpa extract (lower panel). (B) Plot of the HERG current (IHERG) measured at the end of depolarizing pulses against the pulse potential in different concentrations of L. erythrocarpa extract (obtained from A). (C) Plot of the normalized tail current measured at its peak just after repolarization. The amplitude of the tail current in the absence of L. erythrocarpa extract was taken as one. Control data were fitted to the Boltzmann equation, y=1/{1+exp [(-V+V1/2)/dx]}, with V1/2 of -15.2 mV. (D) Activation curves with values normalized to the respective maximum value at each concentration of L. erythrocarpa extract. Symbols with error bars represent means±SEM (n=8).
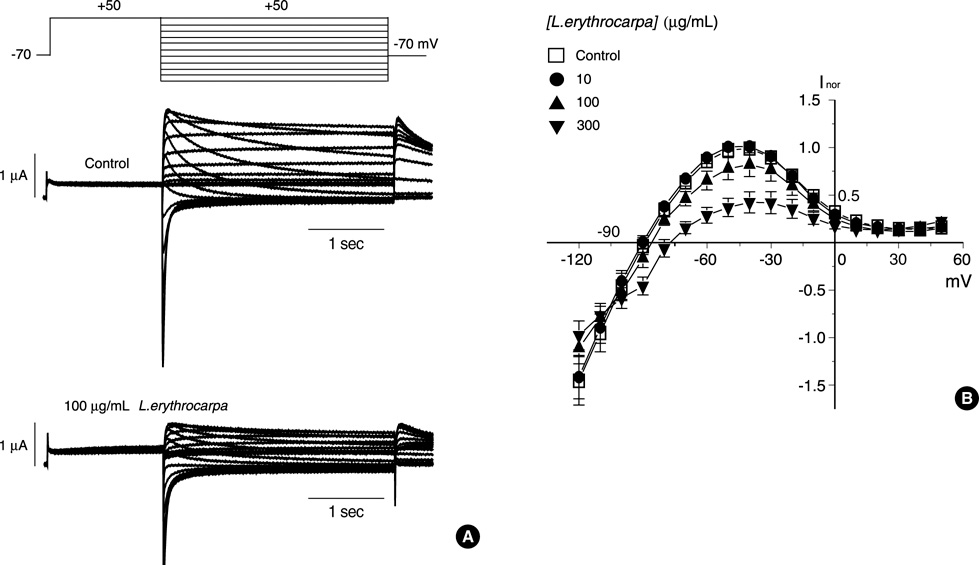
Fig. 2 Effect of L. erythrocarpa extract on the activated current-voltage relationship. (A) Superimposed current traces elicited by various levels of test pulses ranging from -140 to +50 mV following the pre-pulse to +50 mV for 1.5 sec ("inactivation" voltage-clamp protocol, upper panel) before and after the application of 100 µg/mL L. erythrocarpa extract (center and lower panels, respectively). (B) The I-V curve is for the maximal repolarization-evoked (tail) outward currents against the repolarization potential. Symbols with error bars represent the means±S.E.M. (n=9).

Fig. 3 Effect of different solvent fractions of L. erythrocarpa on the HERG current by comparing the changes of IHERG and tail currents with each solvent fraction: the hexane fraction, CHCl3 fraction, EtOAc fraction, BuOH fraction, and H2O fraction at the same concentrations (100 µg/mL). (A) Superimposed current traces from a cell depolarized to +30 mV before and after exposure to the hexane fraction, CHCl3 fraction, EtOAc fraction, BuOH fraction, and H2O fraction of L. erythrocarpa, respectively. (B) Plot of the HERG current (IHERG) measured at the end of depolarizing pulses against the pulse potential in different solvent fractions of L. erythrocarpa (obtained from A). (C) Plot of the normalized tail current measured at its peak just after repolarization. The amplitude of the tail current in the absence of L. erythrocarpa was taken as one. Control data were fitted to the Boltzmann equation, y=1/{1+exp[(-V+V1/2)/dx]}, with V1/2 of -15.3 mV. (D) Activation curves with values normalized to the respective maximum value for each fraction of L. erythrocarpa. Symbols with error bars represent means±SEM (n=8-22).
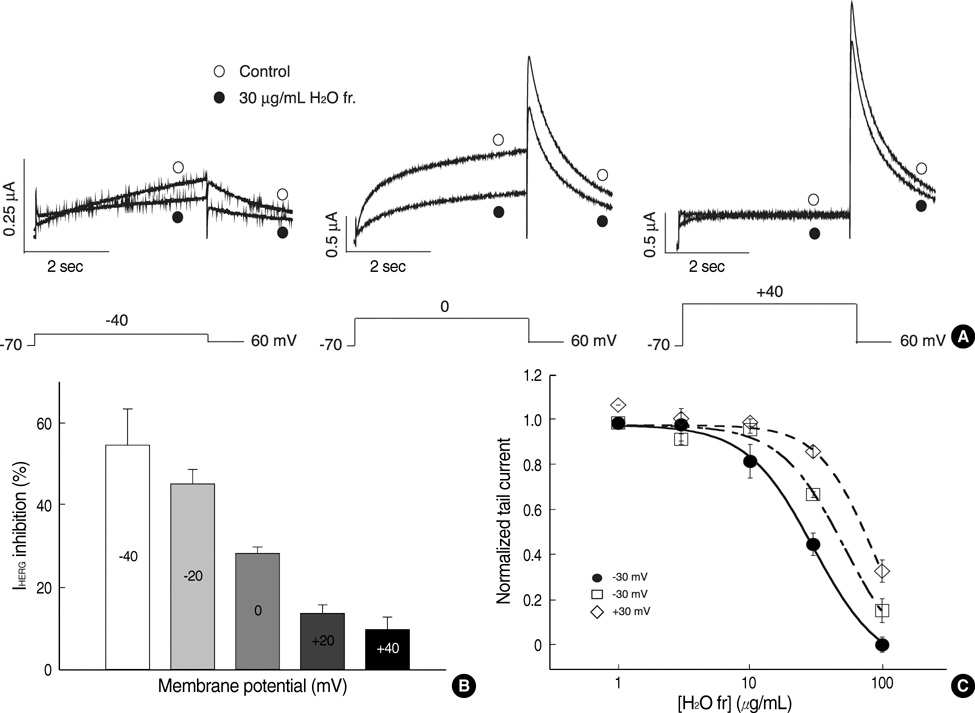
Fig. 4 Voltage dependence of HERG current blocked by the H2O fraction of L. erythrocarpa. (A) Current traces from a cell depolarized to -40 mV (left panel), 0 mV (middle panel) and +40 mV (right panel), before and after exposure to 30 µg/mL of the H2O fraction of L. erythrocarpa, showing increased blockade of HERG current at the more negative potential. The protocol consisted of 4-sec depolarizing steps to -40 mV, 0 mV or +40 mV from a holding potential of -70 mV, followed by repolarization to -60 mV. (B) HERG current inhibition at different voltages by the H2O fraction of L. erythrocarpa. At each depolarizing voltage step (-40, -20, 0, +20 or +40 mV), the tail currents in the presence of 30 µg/mL H2O fraction of L. erythrocarpa were normalized to the tail current obtained in the absence of L. erythrocarpa. Bars with error bars represent means±SEM (n=6). (C) Concentration-dependent block of HERG current by the H2O fraction at different membrane potentials. At each depolarizing voltage step (-30 mV, 0 mV or +30 mV), the tail currents in the presence of various concentrations of the H2O fraction were normalized to the tail current obtained in the absence of L. erythrocarpa, and plotted against H2O fraction concentrations. Symbols with error bars represent means±SEM (n=6). The line represents the data fits to the Hill equation.

Fig. 5 Voltage dependence of HERG current blocked by the BuOH fraction of L. erythrocarpa. (A) Current traces from a cell depolarized to -40 mV (left panel), 0 mV (middle panel) and +40 mV (right panel), before and after exposure to 30 µg/mL of the BuOH fraction of L. erythrocarpa, showing an increased blockade of HERG current at the more negative potential. The protocol consisted of 4-sec depolarizing steps to -40 mV, 0 mV or +40 mV from a holding potential of -70 mV, followed by repolarization to -60 mV. (B) HERG current inhibition at different voltages by the BuOH fraction of L. erythrocarpa. At each depolarizing voltage step (-40, -20, 0, +20 or +40 mV), the tail currents in the presence of 30 µg/mL BuOH fraction of L. erythrocarpa were normalized to the tail current obtained in the absence of L. erythrocarpa. Bars with error bars represent means±SEM (n=6). (C) Concentration-dependent block of HERG current by the BuOH fraction at different membrane potentials. At each depolarizing voltage step (-30 mV, 0 mV or +30 mV), the tail currents in the presence of various concentrations of the BuOH fraction were normalized to the tail current obtained in the absence of L. erythrocarpa, and plotted against BuOH fraction concentrations. Symbols with error bars represent means±SEM (n=6). The line represents the data fits to the Hill equation.
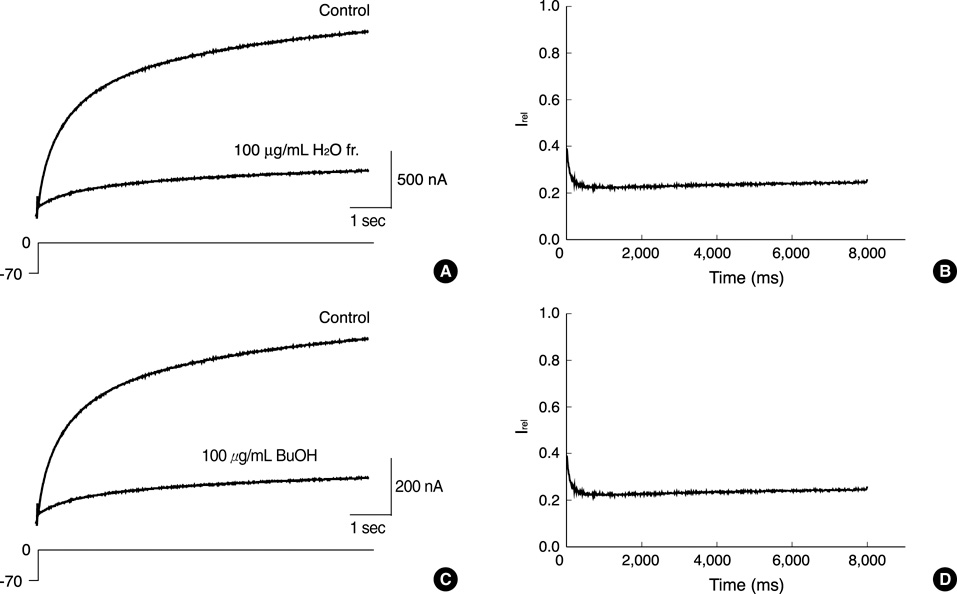
Fig. 6 Relative change in sustained HERG currents in response to H2O or BuOH fractions of L. erythrocarpa. (A, C) Original recording of currents under control conditions (Control) and in the presence of H2O (A) or BuOH (C) fraction of L. erythrocarpa (100 µg/mL each) during voltage steps to 0 mV. After having recorded the control measurement, the oocyte was clamped at -70 mV for 13 min during superfusion with the each fraction. (B, D) Relative current (Irel) obtained by dividing the H2O (C) or BuOH (D) fraction current by the control currents of the recording in A or C, respectively. Time 0 ms corresponds to the beginning of the depolarizing voltage step.
Reference
-
1. Sun BY, Chung YH. Monbographic study of the Lauracea in Korea. Kor J Plant Tax. 1988. 18:133–151.2. Liu SY, Hisada S, Inagaki I. Terpenses of Lindera erythrocarpa. Phytochemistry. 1973. 12:233.3. Liu SY, Hisada S, Inagaki I. Constituents of Lindera erythrocarpa. Phytochemistry. 1973. 12:472.4. Oh HM, Choi SK, Lee JM, Lee SK, Kim HY, Han DC, Kim HM, Son KH, Kwon BM. Cyclopentenediones, inhibitors of farnesyl protein transferase and anti-tumor compounds, isolated from the fruit of Lindera erythrocarpa Makino. Bioorg Med Chem. 2005. 13:6182–6187.5. Lee SM, Baek SH, Lee CH, Lee HB, Kho YH. Cytotoxicity of Lignans from Lindera erythrocarpa Makino. Nat Prod Sci. 2002. 8:100–102.6. Sanguinetti MC, Jurkiewicz NK. Two components of cardiac delayed rectifier K+ current. Differential sensitivity to block by class III anti-arrhythmic agents. J Gen Physiol. 1990. 96:195–215.7. Spector PS, Curran ME, Zou A, Keating MT, Sanguinetti MC. Fast inactivation causes rectification of the IKr channel. J Gen Physiol. 1996. 107:611–619.
Article8. Curran ME, Splawski I, Timothy KW, Vincent GM, Green ED, Keating MT. A molecular basis for cardiac arrhythmia: HERG mutations cause long QT syndrome. Cell. 1995. 80:795–803.
Article9. Bianchi L, Wible B, Arcangeli A, Taglialatela M, Morra F, Castaldo P, Crociani O, Rosati B, Faravelli L, Olivotto M, Wanke E. Herg encodes a K+ current highly conserved in tumors of different histogenesis: a selective advantage for cancer cells? Cancer Res. 1998. 58:815–822.10. Cherubini A, Taddei GL, Crociani O, Paglierani M, Buccoliero AM, Fontana L, Noci I, Borri P, Borrani E, Giachi M, Becchetti A, Rosati B, Wanke E, Olivotto M, Arcangeli A. HERG potassium channels are more frequently expressed in human endometrial cancer as compared to non-cancerous endometrium. Br J Cancer. 2000. 83:1722–1729.
Article11. Pardo LA, del Camino D, Sánchez A, Alves F, Brüggemann A, Beckh S, Stühmer W. Oncogenic potential of EAG K(+) channels. EMBO J. 1999. 18:5540–5547.
Article12. Suzuki T, Takimoto K. Selective expression of HERG and Kv2 channels influences proliferation of uterine cancer cells. Inter J Oncol. 2004. 25:153–159.
Article13. Sanguinetti MC, Jiang C, Curran ME, Keating MT. A mechanistic link between an inherited and an acquired cardiac arrhythmia: HERG encodes the IKr potassium channel. Cell. 1995. 81:299–307.14. Kiehn J, Lacerda AE, Wible B, Brown AM. Molecular physiology and pharmacology of HERG. Single-channel currents and block by dofetilide. Circulation. 1996. 94:2572–2579.15. Thomas D, Wendt-Nordahl G, Rockl K, Ficker E, Brown AM, Kiehn J. High-affinity blockade of human ether-a-go-go-related gene human cardiac potassium channels by the novel antiarrhythmic drug BRL-32872. J Pharmacol Exp Ther. 2001. 297:753–761.16. Parihar AS, Coghlan MJ, Gopalakrishnan M, Shieh CC. Effects of intermediate-conductance Ca2+-activated K+ channel modulators on human prostate cancer cell proliferation. Eur J Pharmacol. 2003. 471:157–164.
Article17. Inglis V, Karpinski E, Benishin C. Gamma-dendrotoxin blocks large conductance Ca2+-activated K+ channels in neuroblastoma cells. Life Sci. 2003. 73:2291–2305.
Article18. Carignani C, Roncarati R, Rimini R, Terstappen GC. Pharmacological and molecular characterisation of SK3 channels in the TE671 human medulloblastoma cell line. Brain Res. 2002. 939:11–18.
Article19. Wu H, Wu K, Han Y, Shi Y, Yao L, Wang J, Fan D. Delayed rectifier K(+) channel regulated by cyclooxygenase-2 in human gastric cancer cell. Zhonghua Zhong Liu Za Zhi. 2002. 24:440–443.20. Soliven B, Ma L, Bae H, Attali B, Sobko A, Iwase T. PDGF upregulates delayed rectifier via Src family kinases and sphingosine kinase in oligodendroglial progenitors. Am J Physiol Cell Physiol. 2003. 284:C85–C93.21. Wang H, Zhang Y, Cao L, Han H, Wang J, Yang B, Nattel S, Wang Z. HERG K+ channel, a regulator of tumor cell apoptosis and proliferation. Cancer Res. 2002. 62:4843–4848.22. Cone CD Jr, Tongier M Jr. Control of somatic cell mitosis by simulated changes in the transmembrane potential level. Oncology. 1971. 25:168–182.
Article23. Rim YS, Park YM, Park MS, Kim KY, Kim MJ, Choi YH. Screening of antioxidants and antimicrobial activity in native plants. Korean J Med Crop Sci. 2000. 8:342–350.24. Suessbrich H, Schonherr R, Heinemann SH, Attali B, Lang F, Busch AE. The inhibitory effect of the antipsychotic drug haloperidol on HERG potassium channels expressed in Xenopus oocytes. Br J Pharmacol. 1997. 120:968–974.
Article25. Suessbrich H, Waldegger S, Lang F, Busch AE. Blockade of HERG channels expressed in Xenopus oocytes by the histamine receptor antagonists terfenadine and astemizole. FEBS Lett. 1996. 385:77–80.
Article26. Rampe D, Roy ML, Dennis A, Brown AM. A mechanism for the proarrhythmic effects of cisapride (Propulsid): high affinity blockade of the human cardiac potassium channel HERG. FEBS Lett. 1997. 417:28–32.
Article27. Green WN, Andersen OS. Surface charges and ion channel function. Annu Rev Physiol. 1991. 53:341–359.
Article28. Hille B. Ionic channels of excitable membranes. 1992. 2nd ed. Sunderland, Mass: Sinauer Associates Inc;459–461.29. Wang N, Minatoguchi S, Arai M, Uno Y, Hashimoto K, Xue-Hai C, Fukuda K, Akao S, Takemura G, Fujiwara H. Lindera strychnifolia is protective against post-ischemic myocardial dysfunction through scavenging hydroxyl radicals and opening the mitochondrial KATP channels in isolated rat hearts. Am J Chin Med. 2004. 32:587–598.
Article30. Su MJ, Nieh YC, Huang HW, Chen CC. Dicentrine, an alpha-adrenoceptor antagonist with sodium and potassium channel blocking activities. Naunyn-Schmiedebergs Arch Pharmacol. 1994. 349:42–49.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Acepromazine inhibits hERG potassium ion channels expressed in human embryonic kidney 293 cells
- Taxifolin Glycoside Blocks Human ether-a-go-go Related Gene K+ Channels
- Effects of Paroxetine on a Human Ether-a-go-go-related Gene (hERG) K⺠Channel Expressed in Xenopus Oocytes and on Cardiac Action Potential
- Block of hERG K+ Channel by Classic Histamine H1 Receptor Antagonist Chlorpheniramine
- Inhibitory Actions of HERG Currents by the Immunosuppressant Drug Cyclosporin A
