Protaetia brevitarsis larvae extract protects against lipopolysaccharidesinduced ferroptosis and inflammation by inhibiting acid sphingomyelinase
- Affiliations
-
- 1Department of Biochemistry, Chung-Ang University College of Medicine, Seoul 06974, Korea
- 2Interdisciplinary Program in Sustainable Living System, Graduate School, Korea University, Seoul 02841, Korea
- 3Department of Human Ecology, Graduate School, Korea University, Seoul 02841, Korea
- KMID: 2560047
- DOI: http://doi.org/10.4162/nrp.2024.18.5.602
Abstract
- BACKGROUND/OBJECTIVES
Inflammation and ferroptosis are implicated in various diseases and lipopolysaccharides (LPS) have been linked with these disorders. Recently, many edible insects, such as Gryllus bimaculatus, Protaetia brevitarsis larvae (PB) and Tenebrio molitor larvae, have been recommended as alternative foods because they contain lots of nutritional sources. In this study, we explored the potential of PB extract in preventing LPS-induced inflammation and ferroptosis in Hep3B cells.
MATERIALS/METHODS
PB powder was extracted using 70% ethanol and applied to Hep3B cells. Co-treatment with LPS was conducted to induce ferroptosis and inflammation. The antiinflammatory and anti-ferroptosis mechanisms of the PB extract were confirmed using Western blot, enzyme-linked immunosorbent assay, and real-time polymerase chain reaction analysis.
RESULTS
PB extract effectively prevented LPS-induced cell death and restored LPS-induced inflammatory cytokine production, NF-κB signaling, endoplasmic reticulum (ER) stress and ferroptosis. Interestingly, PB extract reduced LPS-induced ceramide increase and acid sphingomyelinase (ASMase) expression. The use of the ASMase inhibitor, desipramine, also demonstrated a reduction in these pathways, highlighting the pivotal role of ASMase in inflammation and ferroptosis. Treatment with each inhibitor revealed that ferroptosis causes ER stress and that NF-κB and MAP kinase pathways are involved in inflammation.
CONCLUSION
PB emerges as a potential functional food with inhibitory effects on LPS-induced inflammation and ferroptosis, making it a promising candidate for nutritional interventions.
Figure
-
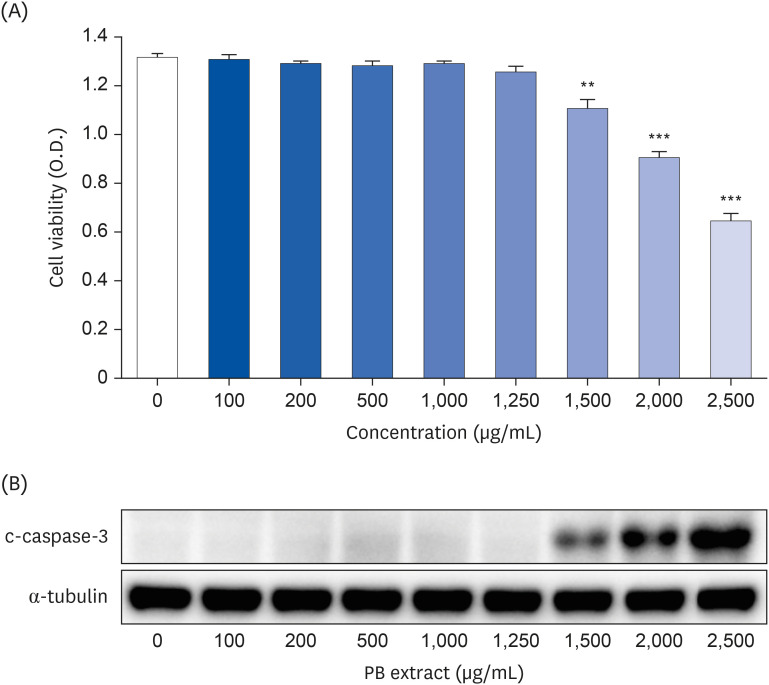
Fig. 1 Effect of PB extract on Hep3B cell death. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with various concentrations of PB extract for 48 h. (A) MTT cell viability assay and (B) representative western blots of c-caspase-3 were examined. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.PB, Protaetia brevitarsis; c-caspase-3, cleaved caspase-3.**P < 0.01, ***P < 0.001.
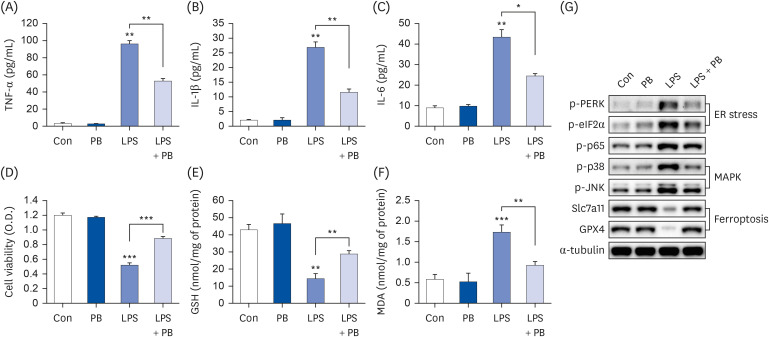
Fig. 2 PB extract reduces LPS-induced inflammation, ferroptosis and cell death. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with PB extract (100 µg/mL) for 24 h, followed by 20 µg/mL LPS for another 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in cell culture media were measured using commercial enzyme-linked immunosorbent assay kits (n = 4). (D) MTT cell viability assay, (E) GSH levels, and (F) MDA levels were examined (n = 4). (G) Representative western blots of indicated proteins. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.TNF, tumor necrosis factor; Con, control (untreated); PB, Protaetia brevitarsis; LPS, lipopolysaccharides; IL, interleukin; GSH, glutathione; MDA, malondialdehyde; PERK, protein kinase R-like endoplasmic reticulum kinase; eIF2α, eukaryotic translation initiation factor 2A; Slc7a11, soluble carrier family 7 member 11; GPX4, glutathione peroxidase 4.*P < 0.05, **P < 0.01, ***P < 0.001.
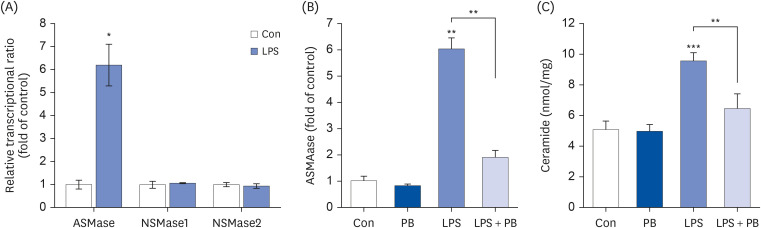
Fig. 3 PB extract reduces LPS-induced acid sphingomyelinase expression. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with PB extract (100 µg/mL) for 24 h, followed by 20 µg/mL LPS for another 24 h. (A) mRNA levels of ASMase, NSMase1, and NSMase2 after 20 µg/mL LPS treatment for 24 h in Hep3B cells. mRNA levels of ASMase (B), and ceramide levels (C) after cotreatment LPS with PB extract, (n = 4). Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.Con, control (untreated); LPS, lipopolysaccharides; ASMase, acid sphingomyelinase; NSMase, neutral sphingomyelinase; PB, Protaetia brevitarsis.*P < 0.05, **P < 0.01, ***P < 0.001.
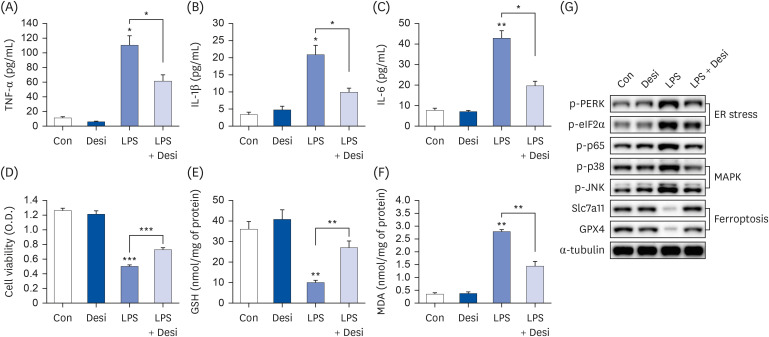
Fig. 4 Desipramine reduces LPS-induced inflammation, ferroptosis and cell death. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with desipramine (2 µM) for 24 h, followed by 20 µg/mL LPS for another 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in cell culture media were measured using commercial enzyme-linked immunosorbent assay kits (n = 4). (D) MTT cell viability assay, (E) GSH levels, and (F) MDA levels were examined (n = 4). (G) Representative western blots of indicated proteins. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.TNF, tumor necrosis factor; Con, control (untreated); Desi, desipramine; LPS, lipopolysaccharides; IL, interleukin; GSH, glutathione; MDA, malondialdehyde; PERK, protein kinase R-like endoplasmic reticulum kinase; eIF2α, eukaryotic translation initiation factor 2A; Slc7a11, soluble carrier family 7 member 11; GPX4, glutathione peroxidase 4; ER, endoplasmic reticulum.*P < 0.05, **P < 0.01, ***P < 0.001.
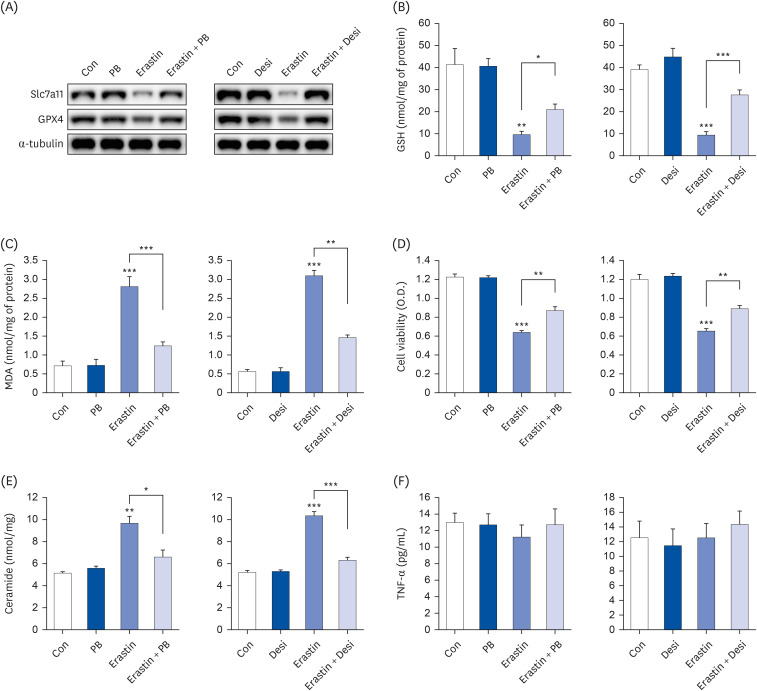
Fig. 5 PB extract and desipramine decrease erastin-induced ferroptosis and cell death. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with PB extract (100 µg/mL) or desipramine (2 µM) for 24 h, followed by 20 µM erastin for another 24 h. (A) Representative western blots of indicated proteins after erastin treatment in PB extract- or desipramine-pretreated Hep3B cells. GSH levels (B), MDA levels (C), MTT cell viability assay (D), ceramide levels (E), and TNF-α (F) were measured after erastin treatment in PB extract- or desipramine-pretreated Hep3B cells (n = 4). Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.Con, control (untreated); PB, Protaetia brevitarsis; Desi, desipramine; Slc7a11, soluble carrier family 7 member 11; GPX4, glutathione peroxidase 4; GSH, glutathione; MDA, malondialdehyde; TNF, tumor necrosis factor.*P < 0.05, **P < 0.01, ***P < 0.001.
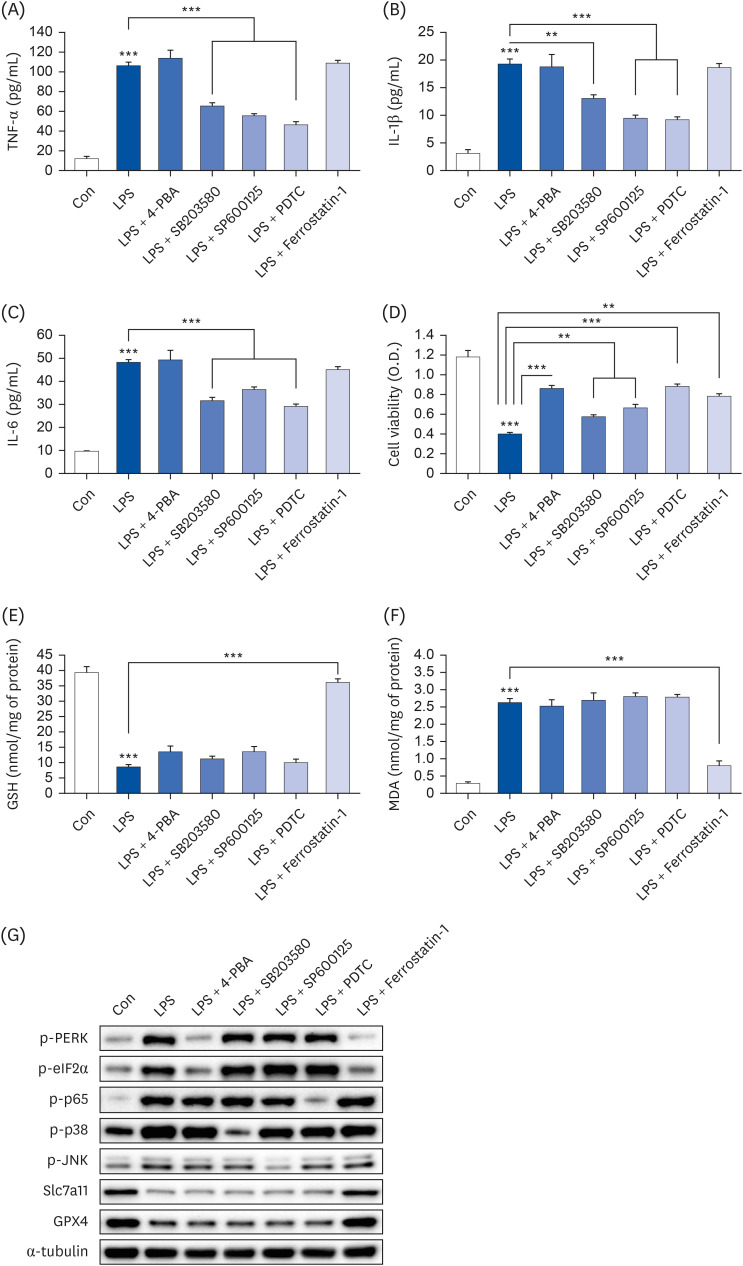
Fig. 6 Ferroptosis/ER stress and NF-κB play an important role in LPS-induced cell death. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with various inhibitors (4-PBA [ER stress inhibitor, 5 mM], SB203580 [p38 inhibitor, 10 µM], SP600125 [JNK inhibitor, 10 µM], PDTC [NF-κB inhibitor, 10 µM], and ferrostatin-1 [ferroptosis inhibitor, 1 µM]) for 24 h, followed by 20 µg/mL LPS for another 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in cell culture media were measured using commercial enzyme-linked immunosorbent assay kits (n = 4). MTT cell viability assay (D), GSH levels (E), and MDA levels (F) were examined (n = 4). (G) Representative western blots of indicated proteins. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.TNF, tumor necrosis factor; Con, control (untreated); LPS, lipopolysaccharides; IL, interleukin; GSH, glutathione; MDA, malondialdehyde; ERK, protein kinase R-like endoplasmic reticulum kinase; eIF2α, eukaryotic translation initiation factor 2A; Slc7a11, soluble carrier family 7 member 11; GPX4, glutathione peroxidase 4; NF, nuclear factor.**P < 0.01, ***P < 0.001.
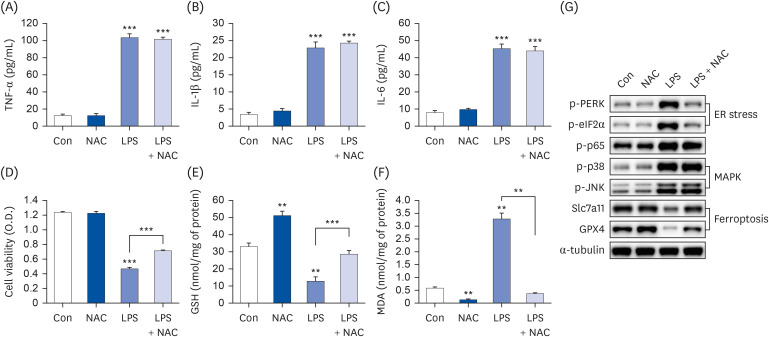
Fig. 7 N-acetylcysteine only inhibits ferroptosis/ER stress, but not inflammatory cytokine production. Hep3B cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with 10 mM NAC for 24 h, followed by 20 µg/mL LPS for another 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in cell culture media were measured using commercial enzyme-linked immunosorbent assay kits (n = 4). MTT cell viability assay (D), GSH levels (E), and MDA levels (F) were examined (n = 4). (G) Representative western blots of indicated proteins. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.TNF, tumor necrosis factor; Con, control (untreated); NAC, N-acetylcysteine; LPS, lipopolysaccharides; IL, interleukin; GSH, glutathione; MDA, malondialdehyde; PERK, protein kinase R-like endoplasmic reticulum kinase; eIF2α, eukaryotic translation initiation factor 2A; GPX4, glutathione peroxidase 4; Slc7a11, soluble carrier family 7 member 11; ER, endoplasmic reticulum.**P < 0.01, ***P < 0.001.
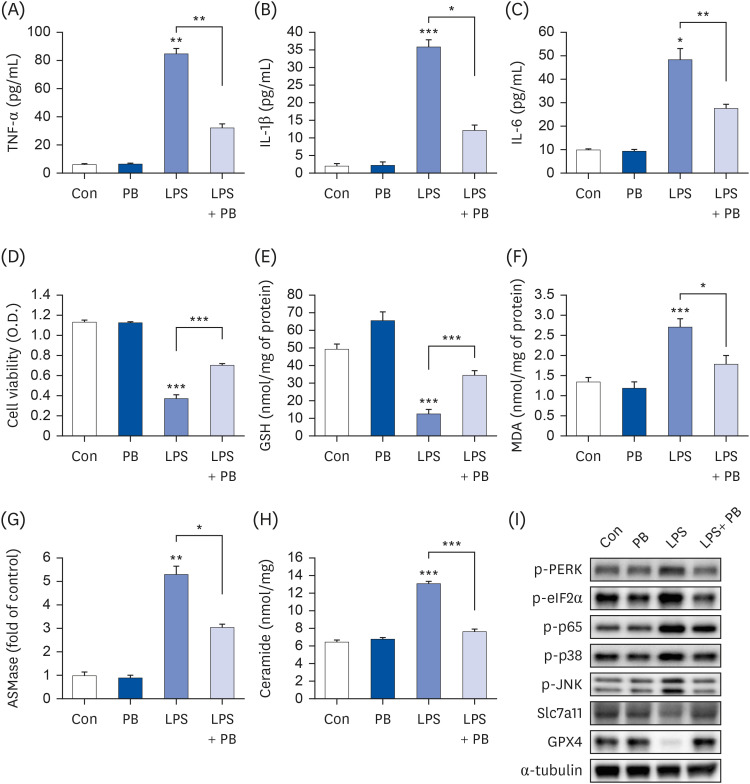
Fig. 8 PB extract reduces LPS-induced inflammation, ferroptosis and cell death in THLE-2 cells. THLE-2 cells were seeded at 3 × 104 cells/well in a 96-well plate and 4 × 105 cells/well in a 6-well plate. They were treated with PB extract (100 µg/mL) for 24 h, followed by 20 µg/mL LPS for another 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in cell culture media were measured using commercial enzyme-linked immunosorbent assay kits (n = 4). (D) MTT cell viability assay, (E) GSH levels, (F) MDA levels, (G) ASMase expression, and (H) ceramide levels were examined (n = 4). (I) Representative western blots of indicated proteins. Values are expressed as mean ± SEM compared to control group. All experiments were performed in triplicate.TNF, tumor necrosis factor; Con, control (untreated); PB, Protaetia brevitarsis; LPS, lipopolysaccharides; IL, interleukin; GSH, glutathione; MDA, malondialdehyde; ASMase, acid sphingomyelinase; PERK, protein kinase R-like endoplasmic reticulum kinase; eIF2α, eukaryotic translation initiation factor 2A; Slc7a11, soluble carrier family 7 member 11; GPX4, glutathione peroxidase 4.*P < 0.05, **P < 0.01, ***P < 0.001.
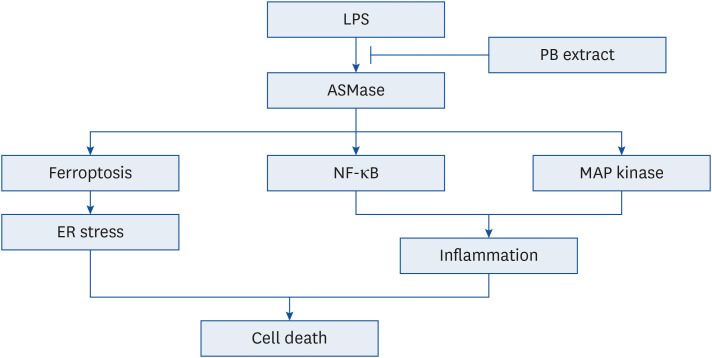
Fig. 9 Schematic showing the effects of PB extract in LPS-induced cell death. LPS induced ASMase activation, initiating a cascade that triggers ferroptosis, NF-κB, and MAP kinase pathways. Ferroptosis promotes ER stress, while NF-κB and MAP kinase pathways contribute to increased inflammation. Both pathways finally lead to cell death. PB extract inhibits ASMase activation, effectively inhibiting all these pathways and preventing cell death.LPS, lipopolysaccharides; PB, Protaetia brevitarsis; ASMase, acid sphingomyelinase; NF, nuclear factor; ER, endoplasmic reticulum.
Reference
-
1. Jantzen da Silva Lucas A, Menegon de Oliveira L, da Rocha M, Prentice C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020; 311:126022. PMID: 31869637.2. Boehm E, Borzekowski D, Ververis E, Lohmann M, Böl GF. Communicating food risk-benefit assessments: edible insects as red meat replacers. Front Nutr. 2021; 8:749696. PMID: 34977112.3. Saeb A, Grundmann SM, Gessner DK, Schuchardt S, Most E, Wen G, Eder K, Ringseis R. Feeding of cuticles from Tenebrio molitor larvae modulates the gut microbiota and attenuates hepatic steatosis in obese Zucker rats. Food Funct. 2022; 13:1421–1436. PMID: 35048923.4. Seo M, Goo TW, Chung MY, Baek M, Hwang JS, Kim MA, Yun EY. Tenebrio molitor larvae inhibit adipogenesis through AMPK and MAPKs signaling in 3T3-L1 adipocytes and obesity in high-fat diet-induced obese mice. Int J Mol Sci. 2017; 18:518. PMID: 28264489.5. Yu JM, Jang JY, Kim HJ, Cho YH, Kim D, Kwon O, Cho YJ, An BJ. Antioxidant capacity and Raw 264.7 macrophage anti-inflammatory effect of the Tenebrio molitor. Korean Journal of Food Preservation. 2016; 23:890–898.6. Ahn EM, Myung NY, Jung HA, Kim SJ. The ameliorative effect of Protaetia brevitarsis larvae in HFD-induced obese mice. Food Sci Biotechnol. 2019; 28:1177–1186. PMID: 31275718.7. Park CE, Lee SO. Nrf2-mediated protective effect of protein hydrolysates from Protaetia brevitarsis larvae against oxidative stress-induced hepatotoxicity. Food Sci Biotechnol. 2023; 32:1561–1571. PMID: 37637846.8. Jang HY, Kim JM, Kim JS, Kim BS, Lee YR, Bae JS. Protaetia brevitarsis extract attenuates RANKL-induced osteoclastogenesis by inhibiting the JNK/NF-κB/PLCγ2 signaling pathway. Nutrients. 2023; 15:3193. PMID: 37513611.9. MacKichan ML, DeFranco AL. Role of ceramide in lipopolysaccharide (LPS)-induced signaling. LPS increases ceramide rather than acting as a structural homolog. J Biol Chem. 1999; 274:1767–1775. PMID: 9880559.10. Xiang H, Jin S, Tan F, Xu Y, Lu Y, Wu T. Physiological functions and therapeutic applications of neutral sphingomyelinase and acid sphingomyelinase. Biomed Pharmacother. 2021; 139:111610. PMID: 33957567.11. García-Ruiz C, Colell A, Marí M, Morales A, Calvo M, Enrich C, Fernández-Checa JC. Defective TNF-alpha-mediated hepatocellular apoptosis and liver damage in acidic sphingomyelinase knockout mice. J Clin Invest. 2003; 111:197–208. PMID: 12531875.12. Fernandez A, Matias N, Fucho R, Ribas V, Von Montfort C, Nuño N, Baulies A, Martinez L, Tarrats N, Mari M, et al. ASMase is required for chronic alcohol induced hepatic endoplasmic reticulum stress and mitochondrial cholesterol loading. J Hepatol. 2013; 59:805–813. PMID: 23707365.13. Lang PA, Schenck M, Nicolay JP, Becker JU, Kempe DS, Lupescu A, Koka S, Eisele K, Klarl BA, Rübben H, et al. Liver cell death and anemia in Wilson disease involve acid sphingomyelinase and ceramide. Nat Med. 2007; 13:164–170. PMID: 17259995.14. Osawa Y, Seki E, Adachi M, Suetsugu A, Ito H, Moriwaki H, Seishima M, Nagaki M. Role of acid sphingomyelinase of Kupffer cells in cholestatic liver injury in mice. Hepatology. 2010; 51:237–245. PMID: 19821528.15. Moles A, Tarrats N, Morales A, Domínguez M, Bataller R, Caballería J, García-Ruiz C, Fernández-Checa JC, Marí M. Acidic sphingomyelinase controls hepatic stellate cell activation and in vivo liver fibrogenesis. Am J Pathol. 2010; 177:1214–1224. PMID: 20651240.16. Mir IH, Thirunavukkarasu C. The relevance of acid sphingomyelinase as a potential target for therapeutic intervention in hepatic disorders: current scenario and anticipated trends. Arch Toxicol. 2023; 97:2069–2087. PMID: 37248308.17. Thayyullathil F, Cheratta AR, Alakkal A, Subburayan K, Pallichankandy S, Hannun YA, Galadari S. Acid sphingomyelinase-dependent autophagic degradation of GPX4 is critical for the execution of ferroptosis. Cell Death Dis. 2021; 12:26. PMID: 33414455.18. Du YX, Zhao YT, Sun YX, Xu AH. Acid sphingomyelinase mediates ferroptosis induced by high glucose via autophagic degradation of GPX4 in type 2 diabetic osteoporosis. Mol Med. 2023; 29:125. PMID: 37710183.19. Xie Y, Hou W, Song X, Yu Y, Huang J, Sun X, Kang R, Tang D. Ferroptosis: process and function. Cell Death Differ. 2016; 23:369–379. PMID: 26794443.20. Yang WS, Kim KJ, Gaschler MM, Patel M, Shchepinov MS, Stockwell BR. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci U S A. 2016; 113:E4966–E4975. PMID: 27506793.21. Hwang D, Goo TW, Yun EY. In Vitro Protective Effect of Paste and Sauce Extract Made with Protaetia brevitarsis Larvae on HepG2 Cells Damaged by Ethanol. Insects. 2020; 11.22. Ganguly K, Dutta SD, Jeong MS, Patel DK, Cho SJ, Lim KT. Naturally-derived protein extract from Gryllus bimaculatus improves antioxidant properties and promotes osteogenic differentiation of hBMSCs. PLoS One. 2021; 16:e0249291. PMID: 34077422.23. Navarro del Hierro J, Gutiérrez-Docio A, Otero P, Reglero G, Martin D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2020; 309:125742. PMID: 31704068.24. Kim MH, Kim SJ, Kim SH, Park WJ, Han JS. Gryllus bimaculatus-containing diets protect against dexamethasone-induced muscle atrophy, but not high-fat diet-induced obesity. Food Sci Nutr. 2023; 11:2787–2797. PMID: 37324877.25. Kim MH, Park JW, Lee EJ, Kim S, Shin SH, Ahn JH, Jung Y, Park I, Park WJ. C16-ceramide and sphingosine 1-phosphate/S1PR2 have opposite effects on cell growth through mTOR signaling pathway regulation. Oncol Rep. 2018; 40:2977–2987. PMID: 30226616.26. Choi RY, Kim IW, Ji M, Paik MJ, Ban EJ, Lee JH, Hwang JS, Kweon H, Seo M. Protaetia brevitarsis seulensis larvae ethanol extract inhibits RANKL-stimulated osteoclastogenesis and ameliorates bone loss in ovariectomized mice. Biomed Pharmacother. 2023; 165:115112. PMID: 37413903.27. Park YM, Noh EM, Lee HY, Shin DY, Lee YH, Kang YG, Na EJ, Kim JH, Yang HJ, Kim MJ, et al. Anti-diabetic effects of Protaetia brevitarsis in pancreatic islets and a murine diabetic model. Eur Rev Med Pharmacol Sci. 2021; 25:7508–7515. PMID: 34919253.28. Zhao C, Xiao C, Feng S, Bai J. Artemisitene alters LPS-induced oxidative stress, inflammation and ferroptosis in liver through Nrf2/HO-1 and NF-kB pathway. Front Pharmacol. 2023; 14:1177542. PMID: 37180725.29. Kim MH, Ahn HK, Lee EJ, Kim SJ, Kim YR, Park JW, Park WJ. Hepatic inflammatory cytokine production can be regulated by modulating sphingomyelinase and ceramide synthase 6. Int J Mol Med. 2017; 39:453–462. PMID: 28035360.30. Yin L, Dai Y, Cui Z, Jiang X, Liu W, Han F, Lin A, Cao J, Liu J. The regulation of cellular apoptosis by the ROS-triggered PERK/EIF2α/chop pathway plays a vital role in bisphenol A-induced male reproductive toxicity. Toxicol Appl Pharmacol. 2017; 314:98–108. PMID: 27894913.31. Sun J, Chen W, Li S, Yang S, Zhang Y, Hu X, Qiu H, Wu J, Xu S, Chu T. Nox4 promotes RANKL-induced autophagy and osteoclastogenesis via activating ROS/PERK/eIF-2α/ATF4 pathway. Front Pharmacol. 2021; 12:751845. PMID: 34650437.32. Rodríguez-Hernández MA, Chapresto-Garzón R, Cadenas M, Navarro-Villarán E, Negrete M, Gómez-Bravo MA, Victor VM, Padillo FJ, Muntané J. Differential effectiveness of tyrosine kinase inhibitors in 2D/3D culture according to cell differentiation, p53 status and mitochondrial respiration in liver cancer cells. Cell Death Dis. 2020; 11:339. PMID: 32382022.33. Jenkins RW, Canals D, Hannun YA. Roles and regulation of secretory and lysosomal acid sphingomyelinase. Cell Signal. 2009; 21:836–846. PMID: 19385042.34. Insausti-Urkia N, Solsona-Vilarrasa E, Garcia-Ruiz C, Fernandez-Checa JC. Sphingomyelinases and liver diseases. Biomolecules. 2020; 10:1497. PMID: 33143193.35. Oh E, Park WJ, Kim Y. Effects of Tenebrio molitor larvae and its protein derivatives on the antioxidant and anti-inflammatory capacities of tofu. Food Biosci. 2022; 50:102105.36. Park WJ, Han JS. Gryllus bimaculatus extract protects against lipopolysaccharide and palmitate-induced production of proinflammatory cytokines and inflammasome formation. Mol Med Rep. 2021; 23:206. PMID: 33495809.37. Choi YJ, Bae IY. White-spotted flower chafer (Protaetia brevitarsis) ameliorates inflammatory responses in LPS-stimulated RAW 264.7 macrophages. J Insects Food Feed. 2023; 9:1037–1046.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Soil Application of Metarhizium anisopliae JEF-314 Granules to Control, Flower Chafer Beetle, Protaetia brevitarsis seulensis
- Inhibition of Fatty Acid β-Oxidation by Fatty Acid Binding Protein 4 Induces Ferroptosis in HK2 Cells Under High Glucose Conditions
- Salidroside attenuates sepsis-induced acute lung injury by inhibiting ferroptosis-dependent pathway
- Novel non-apoptotic cell death: ferroptosis
- Ferroptosis and its role in gastric and colorectal cancers
