Inhibition of Fatty Acid β-Oxidation by Fatty Acid Binding Protein 4 Induces Ferroptosis in HK2 Cells Under High Glucose Conditions
- Affiliations
-
- 1Department of Nephrology, Kidney and Urology Center, The Seventh Affiliated Hospital, Sun Yat-sen University, China
- 2Renal Division, Peking University Shenzhen Hospital, Shenzhen, China
- KMID: 2541879
- DOI: http://doi.org/10.3803/EnM.2022.1604
Abstract
- Background
Ferroptosis, which is caused by an iron-dependent accumulation of lipid hydroperoxides, is a type of cell death linked to diabetic kidney disease (DKD). Previous research has shown that fatty acid binding protein 4 (FABP4) is involved in the regulation of ferroptosis in diabetic retinopathy. The present study was constructed to explore the role of FABP4 in the regulation of ferroptosis in DKD.
Methods
We first detected the expression of FABP4 and proteins related to ferroptosis in renal biopsies of patients with DKD. Then, we used a FABP4 inhibitor and small interfering RNA to investigate the role of FABP4 in ferroptosis induced by high glucose in human renal proximal tubular epithelial (HG-HK2) cells.
Results
In kidney biopsies of DKD patients, the expression of FABP4 was elevated, whereas carnitine palmitoyltransferase-1A (CP-T1A), glutathione peroxidase 4, ferritin heavy chain, and ferritin light chain showed reduced expression. In HG-HK2 cells, the induction of ferroptosis was accompanied by an increase in FABP4. Inhibition of FABP4 in HG-HK2 cells changed the redox state, sup-pressing the production of reactive oxygen species, ferrous iron (Fe2+), and malondialdehyde, increasing superoxide dismutase, and reversing ferroptosis-associated mitochondrial damage. The inhibition of FABP4 also increased the expression of CPT1A, reversed lipid deposition, and restored impaired fatty acid β-oxidation. In addition, the inhibition of CPT1A could induce ferroptosis in HK2 cells.
Conclusion
Our results suggest that FABP4 mediates ferroptosis in HG-HK2 cells by inhibiting fatty acid β-oxidation.
Keyword
Figure
-
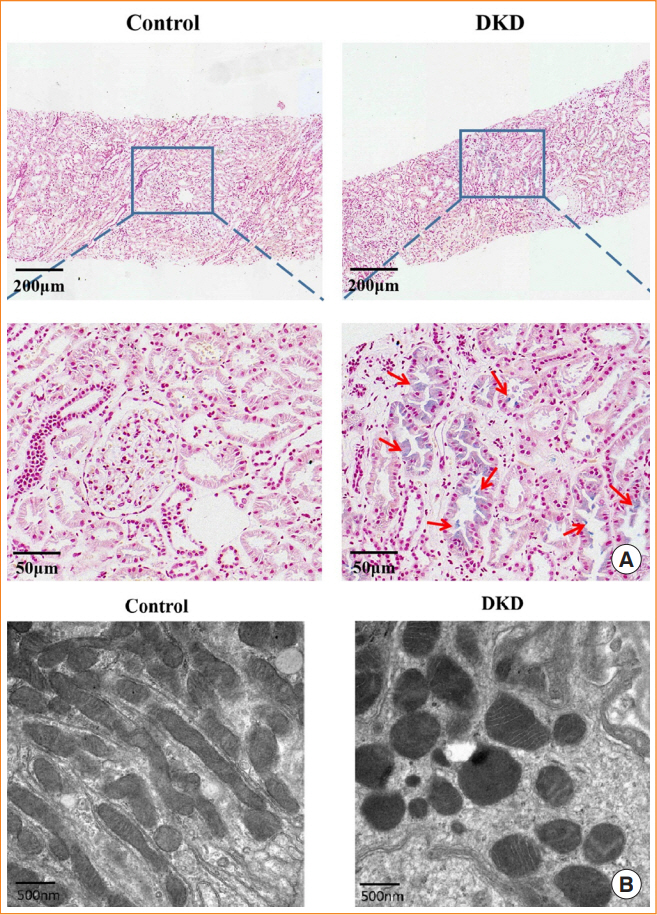
Fig. 1. Changes related to ferroptosis in biopsies of diabetic kidney disease (DKD) patients. (A) Prussian blue staining of kidney biopsy tissue. The red arrows indicate iron deposition. Control refers to patients with glomerular minor lesion. (B) Transmission electron microscopy images of mitochondria in renal proximal tubular epithelial cells. Control refers to patients with early-stage DKD.
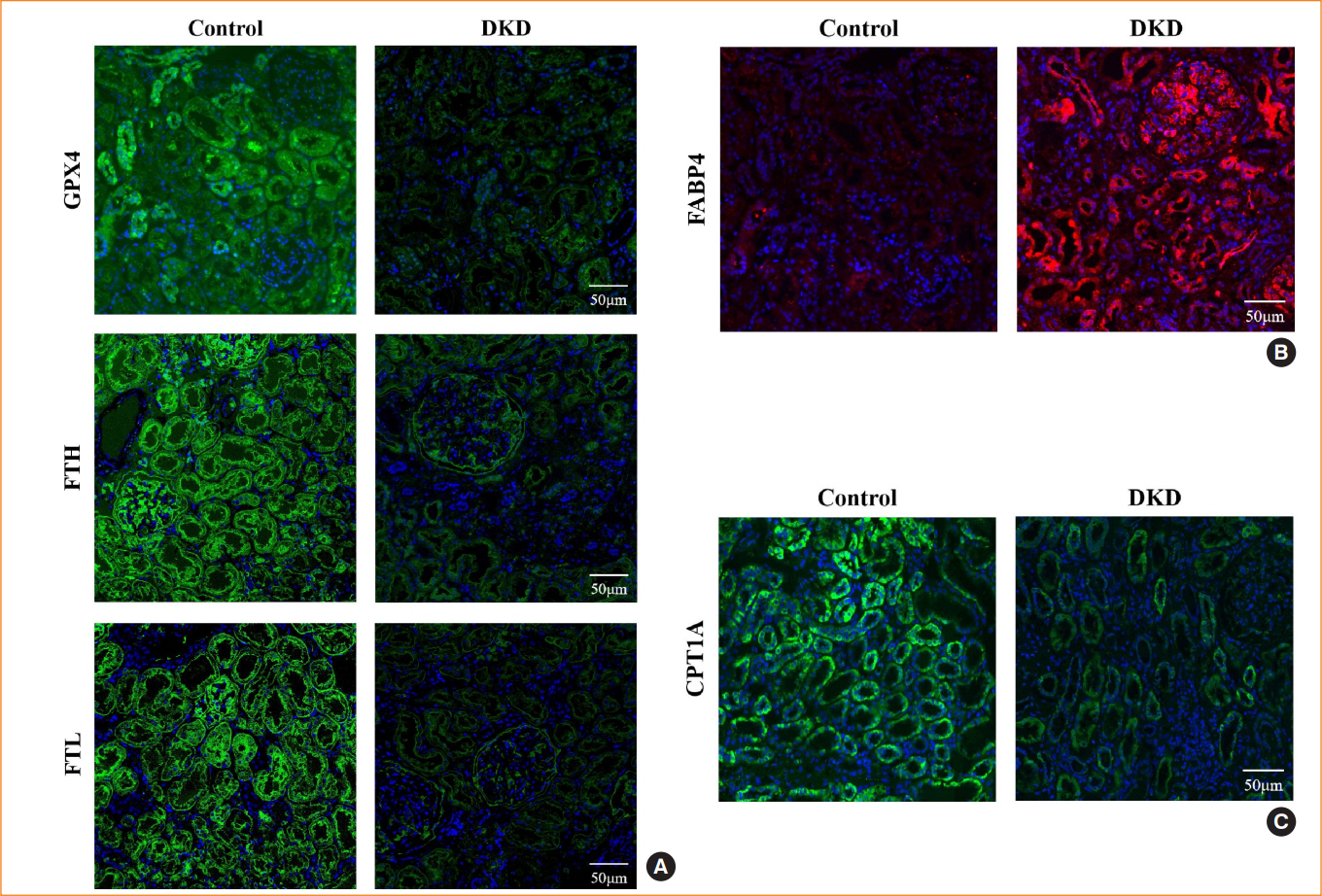
Fig. 2. Expression of fatty acid binding protein 4 (FABP4) and ferroptosis-associated proteins in diabetic kidney disease (DKD) patients. Representative immunofluorescence images of kidney tissue sections in negative controls and DKD patients for (A) glutathione peroxidase 4 (GPX4), ferritin heavy chain (FTH), and ferritin light chain (FTL), (B) FABP4, and (C) carnitine palmitoyltransferase-1A (CPT1A). 4′,6-Diamidino-2-phenylindole (DAPI) indicates nuclear staining.

Fig. 3. Ferroptosis-related changes induced by high glucose in human renal proximal tubular epithelial (HG-HK2) cells. (A) Viability of HK2 cells detected by cell counting kit-8 (CCK-8) in the three groups indicated. Levels of (B) ferrous iron (Fe2+), (C) malondialdehyde (MDA), (D) superoxide dismutase (SOD), and (E) glutathione (GSH) in the control, HG, and HG+ferrostatin-1 (Fer-1) groups. Protein expression of (F) glutathione peroxidase 4 (GPX4), (G) ferritin heavy chain (FTH), ferritin light chain (FTL), and (H) phospho-AMP-activated protein kinase (p-AMPK) in the three groups. (I) Intracellular reactive oxygen species (ROS) production stained with DCFDA/H2DCFDA fluorescent probes. (J) Mitochondrial morphology detected by transmission electron microscopy. The red arrows indicate the damaged mitochondria (mitochondria cristae vanish and membrane rupture). NS, no statistical significance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CTL, control. aP<0.05, bP<0.01, cP<0.001 when compared with the control group; dP<0.05, eP<0.01, fP<0.001 when compared with the HG group.
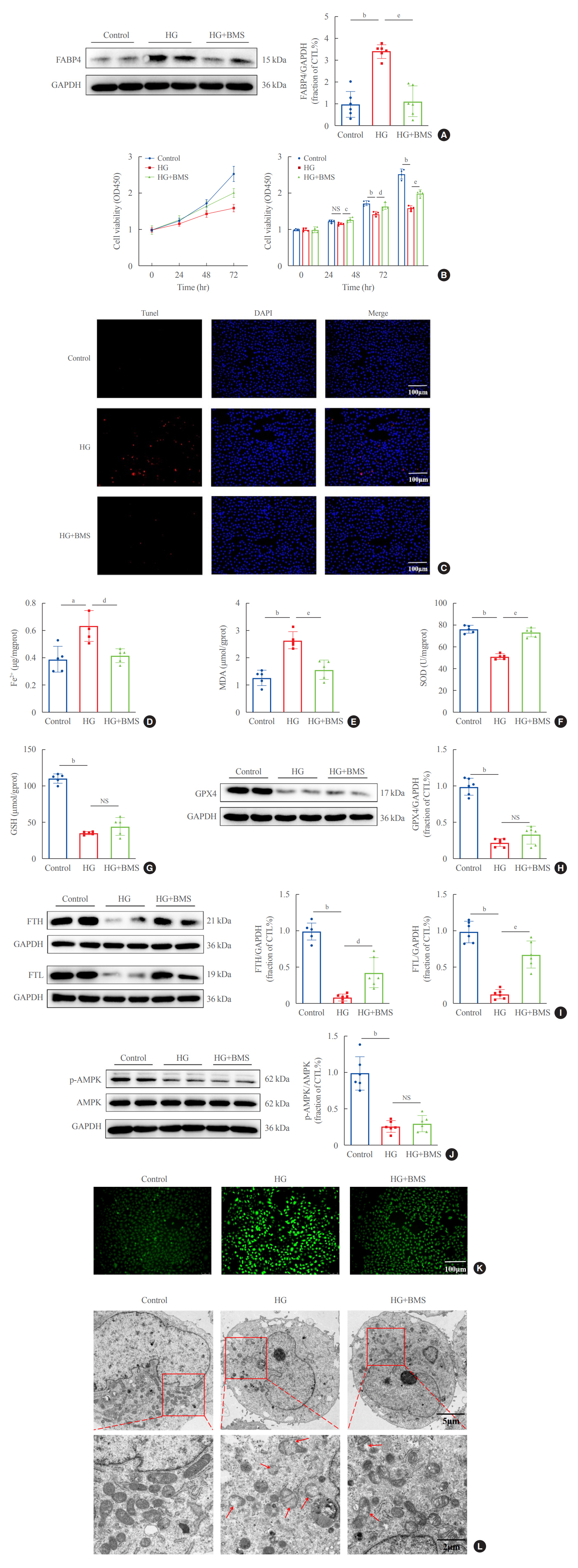
Fig. 4. Inhibition of fatty acid binding protein 4 (FABP4) by BMS309403 (BMS) attenuated ferroptosis induced by high glucose in human renal proximal tubular epithelial (HG-HK2) cells. (A) Expression of FABP4 in the three groups indicated. (B) Viability of cells in the three groups indicated. (C) Representative images of cell death measured by terminal deoxynucleotidyl transferase dUTP nick end labeling (Tunel) staining (positive=red), where the nucleus is labeled with 4′,6-diamidino-2-phenylindole (DAPI; blue). Concentrations of (D) ferrous iron (Fe2+), (E) malondialdehyde (MDA), (F) superoxide dismutase (SOD), (G) glutathione (GSH), (H) expression of glutathione peroxidase 4 (GPX4), (I) ferritin heavy chain (FTH), ferritin light chain (FTL), and (J) phospho-AMP-activated protein kinase (p-AMPK) in each group. (K) Content of reactive oxygen species (ROS) stained with DCFDA/H2DCFDA fluorescent probes. (L) Mitochondrial morphology detected by transmission electron microscopy. The red arrows indicate the damaged mitochondria (mitochondria cristae vanish and membrane rupture). NS, no statistical significance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CTL, control. aP<0.01, bP<0.001 when compared with the control group; cP<0.05, dP<0.01, eP<0.001 when compared with the HG group.

Fig. 5. Silencing fatty acid binding protein 4 (FABP4) inhibited ferroptosis induced by high glucose in human renal proximal tubular epithelial (HG-HK2) cells. (A) Expression of FABP4 after transfection with FABP4-small interfering RNA (siRNA). (B) Effect of silencing FABP4 on cell viability. Concentrations of (C) ferrous iron (Fe2+), (D) malondialdehyde (MDA), (E) superoxide dismutase (SOD), and (F) glutathione (GSH). (G) Reactive oxygen species (ROS) stained with DCFDA/H2DCFDA fluorescent probes. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, no statistical significance. aP<0.001 compared with the negative control (NC)-siRNA group; bP<0.01, cP<0.001 compared with the HG+NC-siRNA group.
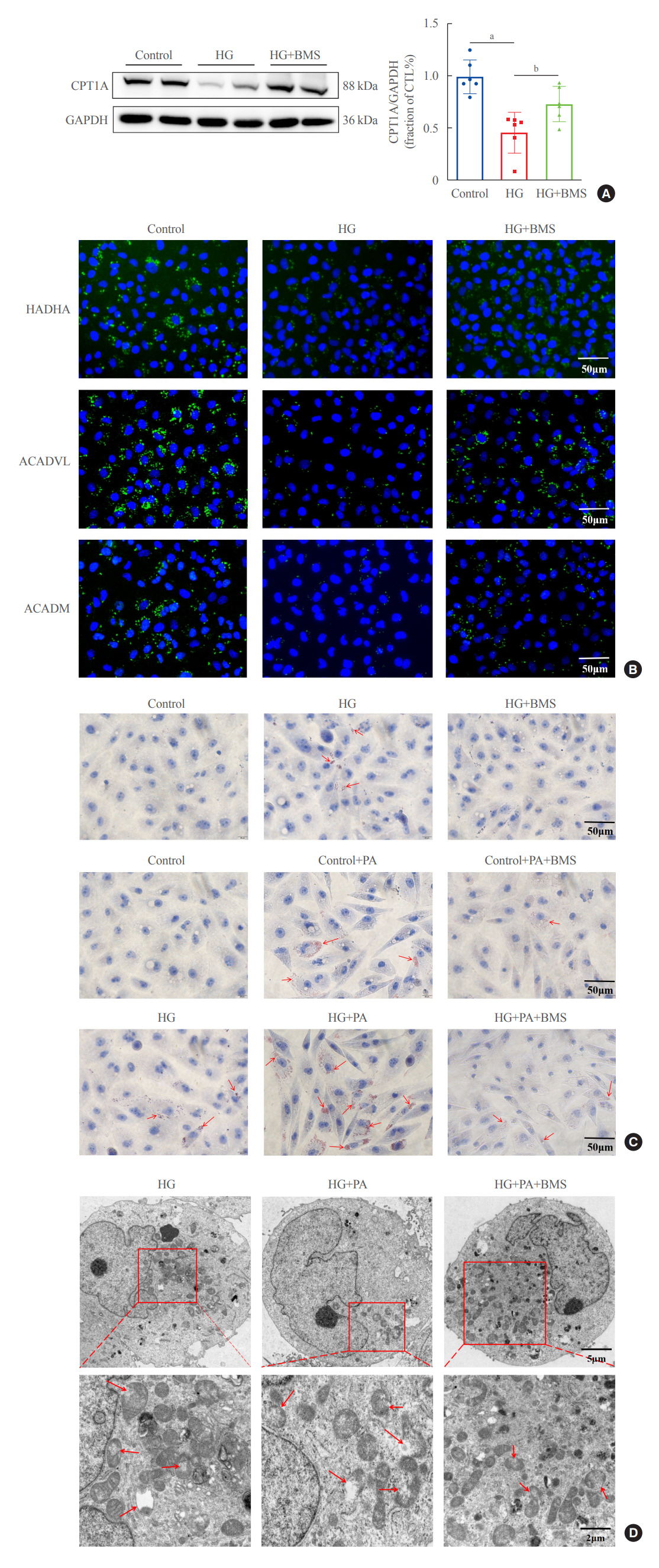
Fig. 6. Inhibition of fatty acid binding protein 4 (FABP4) restores impaired fatty acid β-oxidation in human renal proximal tubular epithelial (HK2) cells. (A) Protein expression of carnitine palmitoyltransferase-1A (CPT1A) in HK2 cells for the three groups indicated. (B) Representative immunofluorescence images of HK2 cells for long chain 3-hydroxyl-coenzyme A (CoA) dehydrogenase (HADHA), acyl-CoA dehydrogenase very long chain (ACADVL), and acyl-CoA dehydrogenase, medium chain specific (ACADM). (C) Lipid accumulation after high glucose (HG) and palmitic acid (PA) stimulation, with and without BMS309403 (BMS), as detected by Oil Red O staining. The red arrows indicate lipid droplets. (D) Mitochondrial morphology in HG, HG+PA, and HG+PA+BMS groups. The red arrows indicate the damaged mitochondria (mitochondria cristae vanish and membrane rupture). GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CTL, control. aP<0.001 compared with the control group; bP<0.05 compared with the HG group.
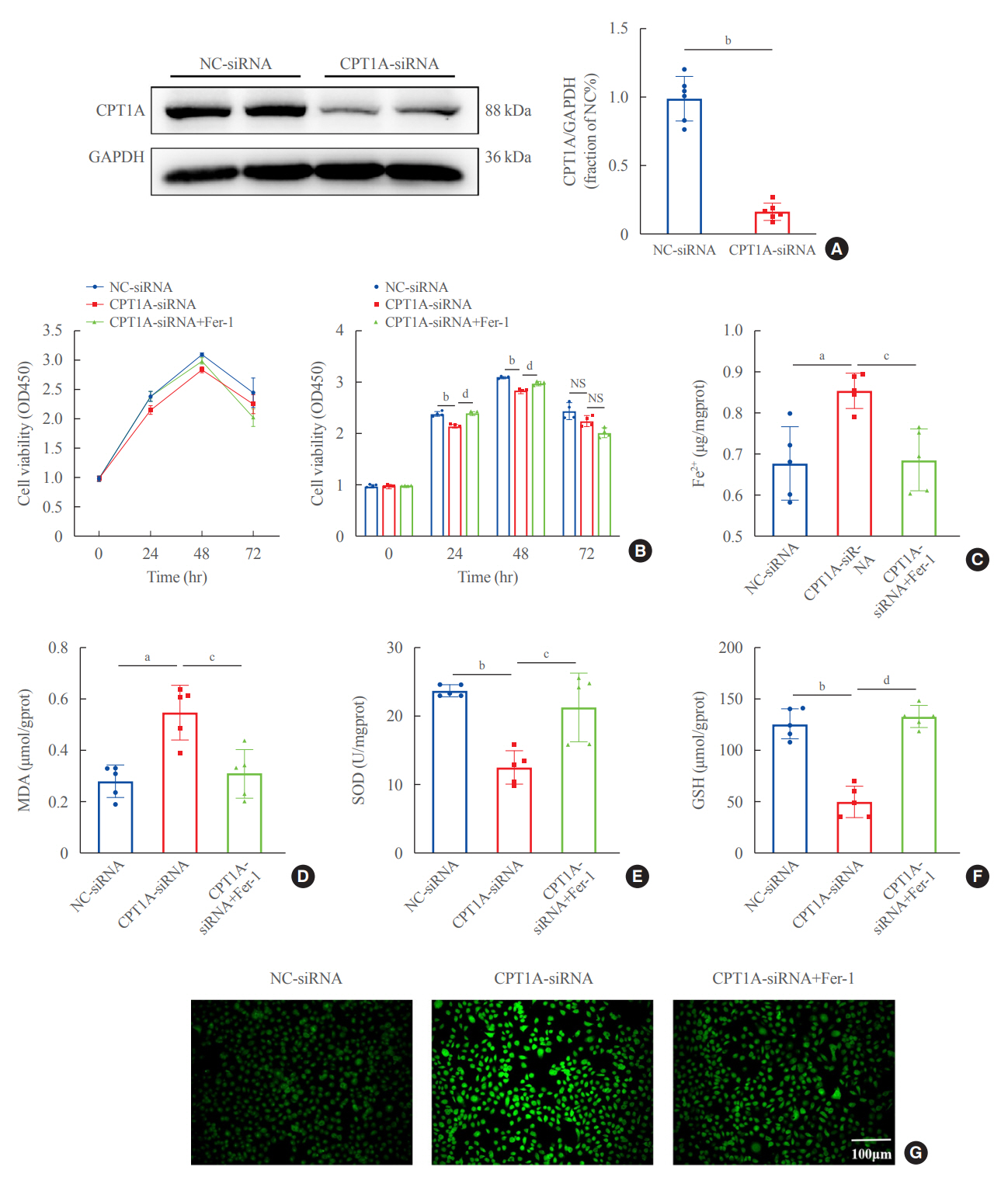
Fig. 7. Silencing carnitine palmitoyltransferase-1A (CPT1A) induced ferroptosis in human renal proximal tubular epithelial (HK2) cells. (A) Expression of CPT1A after transfection with CPT1A-small interfering RNA (siRNA). (B) Effect of silencing CPT1A on cell viability. Concentrations of (C) ferrous iron (Fe2+), (D) malondialdehyde (MDA), (E) superoxide dismutase (SOD), and (F) glutathione (GSH). (G) Reactive oxygen species (ROS) content stained with DCFDA/H2DCFDA fluorescent probes. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Fer-1, ferrostatin-1; NS, no statistical significance. aP<0.01, bP<0.001 compared with the negative control (NC)-siRNA group; cP<0.01, dP<0.001 compared with the CPT1A-siRNA group.

Fig. 8. Potential mechanism of fatty acid binding protein 4 (FABP4) involvement in ferroptosis-mediated renal tubule injury induced by high glucose. Physiologically, fatty acids are taken up from the basolateral membrane by proximal renal tubular epithelial cells through the cluster of differentiation 36 (CD36) receptor; and from the apical membrane through endocytosis in albumin-bound form. In the cell, they are converted to fatty acyl-coenzyme A (CoA) and are transported to mitochondria for fatty acid β-oxidation (FAO). In diabetes (red arrow), high glucose upregulates FABP4 expression in renal tubular cells, which inhibits FAO through downregulating carnitine palmitoyltransferase-1 (CPT1) and leads to lipid accumulation, providing more substrate for lipid peroxidation. Polyunsaturated fatty acids (PUFAs) are converted to PUFA-CoA and are incorporated into PUFA-phospholipids, which react with iron-dependent reactive oxygen species (ROS) and induce lipid peroxidation and ferroptosis. ALB, albumin; FA, fatty acid; Fe3+, ferric iron; TfR1, transferrin receptor 1; TG, triglycerides; Fe2+, ferrous iron; O2−, superoxide anion; SOD, superoxide dismutase; H2O2, hydrogen peroxide; PL, phospholipid.
Reference
-
1. Bonner R, Albajrami O, Hudspeth J, Upadhyay A. Diabetic kidney disease. Prim Care. 2020; 47:645–59.
Article2. Cheng HT, Xu X, Lim PS, Hung KY. Worldwide epidemiology of diabetes-related end-stage renal disease, 2000-2015. Diabetes Care. 2021; 44:89–97.
Article3. Levin A, Tonelli M, Bonventre J, Coresh J, Donner JA, Fogo AB, et al. Global kidney health 2017 and beyond: a roadmap for closing gaps in care, research, and policy. Lancet. 2017; 390:1888–917.
Article4. Mou Y, Wang J, Wu J, He D, Zhang C, Duan C, et al. Ferroptosis, a new form of cell death: opportunities and challenges in cancer. J Hematol Oncol. 2019; 12:34.
Article5. Tang S, Xiao X. Ferroptosis and kidney diseases. Int Urol Nephrol. 2020; 52:497–503.
Article6. Weiland A, Wang Y, Wu W, Lan X, Han X, Li Q, et al. Ferroptosis and its role in diverse brain diseases. Mol Neurobiol. 2019; 56:4880–93.
Article7. Dixon SJ, Lemberg KM, Lamprecht MR, Skouta R, Zaitsev EM, Gleason CE, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012; 149:1060–72.
Article8. Li J, Cao F, Yin HL, Huang ZJ, Lin ZT, Mao N, et al. Ferroptosis: past, present and future. Cell Death Dis. 2020; 11:88.
Article9. Hu W, Liang K, Zhu H, Zhao C, Hu H, Yin S. Ferroptosis and its role in chronic diseases. Cells. 2022; 11:2040.
Article10. Yang WS, Stockwell BR. Ferroptosis: death by lipid peroxidation. Trends Cell Biol. 2016; 26:165–76.
Article11. Ayala A, Munoz MF, Arguelles S. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid Med Cell Longev. 2014; 2014:360438.
Article12. Ursini F, Maiorino M. Lipid peroxidation and ferroptosis: the role of GSH and GPx4. Free Radic Biol Med. 2020; 152:175–85.
Article13. Wang Y, Qiu S, Wang H, Cui J, Tian X, Miao Y, et al. Transcriptional repression of ferritin light chain increases ferroptosis sensitivity in lung adenocarcinoma. Front Cell Dev Biol. 2021; 9:719187.
Article14. He J, Li Z, Xia P, Shi A, FuChen X, Zhang J, et al. Ferroptosis and ferritinophagy in diabetes complications. Mol Metab. 2022; 60:101470.
Article15. Thevenod F, Wolff NA. Iron transport in the kidney: implications for physiology and cadmium nephrotoxicity. Metallomics. 2016; 8:17–42.
Article16. Agborbesong E, Li LX, Li L, Li X. Molecular mechanisms of epigenetic regulation, inflammation, and cell death in ADPKD. Front Mol Biosci. 2022; 9:922428.
Article17. Li S, Zheng L, Zhang J, Liu X, Wu Z. Inhibition of ferroptosis by up-regulating Nrf2 delayed the progression of diabetic nephropathy. Free Radic Biol Med. 2021; 162:435–49.
Article18. Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021; 22:266–82.
Article19. Zheng J, Conrad M. The metabolic underpinnings of ferroptosis. Cell Metab. 2020; 32:920–37.
Article20. Lee JY, Kim WK, Bae KH, Lee SC, Lee EW. Lipid metabolism and ferroptosis. Biology (Basel). 2021; 10:184.
Article21. Lin Z, Liu J, Kang R, Yang M, Tang D. Lipid metabolism in ferroptosis. Adv Biol (Weinh). 2021; 5:e2100396.
Article22. Miess H, Dankworth B, Gouw AM, Rosenfeldt M, Schmitz W, Jiang M, et al. The glutathione redox system is essential to prevent ferroptosis caused by impaired lipid metabolism in clear cell renal cell carcinoma. Oncogene. 2018; 37:5435–50.
Article23. Chen X, Li J, Kang R, Klionsky DJ, Tang D. Ferroptosis: machinery and regulation. Autophagy. 2021; 17:2054–81.
Article24. Opazo-Rios L, Mas S, Marin-Royo G, Mezzano S, Gomez-Guerrero C, Moreno JA, et al. Lipotoxicity and diabetic nephropathy: novel mechanistic insights and therapeutic opportunities. Int J Mol Sci. 2020; 21:2632.
Article25. Russo GT, De Cosmo S, Viazzi F, Pacilli A, Ceriello A, Genovese S, et al. Plasma triglycerides and HDL-C levels predict the development of diabetic kidney disease in subjects with type 2 diabetes: the AMD Annals Initiative. Diabetes Care. 2016; 39:2278–87.
Article26. Herman-Edelstein M, Scherzer P, Tobar A, Levi M, Gafter U. Altered renal lipid metabolism and renal lipid accumulation in human diabetic nephropathy. J Lipid Res. 2014; 55:561–72.
Article27. Saudek CD, Eder HA. Lipid metabolism in diabetes mellitus. Am J Med. 1979; 66:843–52.
Article28. Li H, Xiao Y, Tang L, Zhong F, Huang G, Xu JM, et al. Adipocyte fatty acid-binding protein promotes palmitate-induced mitochondrial dysfunction and apoptosis in macrophages. Front Immunol. 2018; 9:81.
Article29. Li HL, Wu X, Xu A, Hoo RL. A-FABP in metabolic diseases and the therapeutic implications: an update. Int J Mol Sci. 2021; 22:9386.
Article30. Shi M, Ma L, Fu P. Role of fatty acid binding protein 4 (FABP4) in kidney disease. Curr Med Chem. 2020; 27:3657–64.
Article31. Chen Y, Dai Y, Song K, Huang Y, Zhang L, Zhang C, et al. Pre-emptive pharmacological inhibition of fatty acid-binding protein 4 attenuates kidney fibrosis by reprogramming tubular lipid metabolism. Cell Death Dis. 2021; 12:572.
Article32. Tanaka M, Furuhashi M, Okazaki Y, Mita T, Fuseya T, Ohno K, et al. Ectopic expression of fatty acid-binding protein 4 in the glomerulus is associated with proteinuria and renal dysfunction. Nephron Clin Pract. 2014; 128:345–51.
Article33. Hotamisligil GS, Bernlohr DA. Metabolic functions of FABPs: mechanisms and therapeutic implications. Nat Rev Endocrinol. 2015; 11:592–605.
Article34. Ni X, Gu Y, Yu H, Wang S, Chen Y, Wang X, et al. Serum adipocyte fatty acid-binding protein 4 levels are independently associated with radioisotope glomerular filtration rate in type 2 diabetic patients with early diabetic nephropathy. Biomed Res Int. 2018; 2018:4578140.
Article35. Lee CH, Cheung CY, Woo YC, Lui DT, Yuen MM, Fong CH, et al. Prospective associations of circulating adipocyte fatty acid-binding protein levels with risks of renal outcomes and mortality in type 2 diabetes. Diabetologia. 2019; 62:169–77.
Article36. Fan X, Xu M, Ren Q, Fan Y, Liu B, Chen J, et al. Downregulation of fatty acid binding protein 4 alleviates lipid peroxidation and oxidative stress in diabetic retinopathy by regulating peroxisome proliferator-activated receptor γ-mediated ferroptosis. Bioengineered. 2022; 13:10540–51.
Article37. Liu J, Huang R, Li X, Guo F, Li L, Zeng X, et al. Genetic inhibition of FABP4 attenuated endoplasmic reticulum stress and mitochondrial dysfunction in rhabdomyolysis-induced acute kidney injury. Life Sci. 2021; 268:119023.
Article38. Luis G, Godfroid A, Nishiumi S, Cimino J, Blacher S, Maquoi E, et al. Tumor resistance to ferroptosis driven by stearoyl-CoA desaturase-1 (SCD1) in cancer cells and fatty acid biding protein-4 (FABP4) in tumor microenvironment promote tumor recurrence. Redox Biol. 2021; 43:102006.
Article39. Fang X, Ardehali H, Min J, Wang F. The molecular and metabolic landscape of iron and ferroptosis in cardiovascular disease. Nat Rev Cardiol. 2023; 20:7–23.
Article40. Chen X, Kang R, Kroemer G, Tang D. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021; 18:280–96.
Article41. Stockwell BR. Ferroptosis turns 10: emerging mechanisms, physiological functions, and therapeutic applications. Cell. 2022; 185:2401–21.
Article42. Liang D, Minikes AM, Jiang X. Ferroptosis at the intersection of lipid metabolism and cellular signaling. Mol Cell. 2022; 82:2215–27.
Article43. Yang WS, Kim KJ, Gaschler MM, Patel M, Shchepinov MS, Stockwell BR. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci U S A. 2016; 113:E4966–75.
Article44. Magtanong L, Ko PJ, To M, Cao JY, Forcina GC, Tarangelo A, et al. Exogenous monounsaturated fatty acids promote a ferroptosis-resistant cell state. Cell Chem Biol. 2019; 26:420–32.
Article45. Lee H, Zandkarimi F, Zhang Y, Meena JK, Kim J, Zhuang L, et al. Energy-stress-mediated AMPK activation inhibits ferroptosis. Nat Cell Biol. 2020; 22:225–34.
Article46. Juszczak F, Caron N, Mathew AV, Decleves AE. Critical role for AMPK in metabolic disease-induced chronic kidney disease. Int J Mol Sci. 2020; 21:7994.
Article47. Entezari M, Hashemi D, Taheriazam A, Zabolian A, Mohammadi S, Fakhri F, et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: a pre-clinical and clinical investigation. Biomed Pharmacother. 2022; 146:112563.48. Gewin LS. Sugar or fat?: renal tubular metabolism reviewed in health and disease. Nutrients. 2021; 13:1580.
Article49. Thongnak L, Pongchaidecha A, Lungkaphin A. Renal lipid metabolism and lipotoxicity in diabetes. Am J Med Sci. 2020; 359:84–99.
Article50. Stadler K, Goldberg IJ, Susztak K. The evolving understanding of the contribution of lipid metabolism to diabetic kidney disease. Curr Diab Rep. 2015; 15:40.
Article51. Miguel V, Tituana J, Herrero JI, Herrero L, Serra D, Cuevas P, et al. Renal tubule Cpt1a overexpression protects from kidney fibrosis by restoring mitochondrial homeostasis. J Clin Invest. 2021; 131:e140695.
Article52. Shen Y, Chen W, Han L, Bian Q, Fan J, Cao Z, et al. VEGF-B antibody and interleukin-22 fusion protein ameliorates diabetic nephropathy through inhibiting lipid accumulation and inflammatory responses. Acta Pharm Sin B. 2021; 11:127–42.
Article53. Fu Y, Sun Y, Wang M, Hou Y, Huang W, Zhou D, et al. Elevation of JAML promotes diabetic kidney disease by modulating podocyte lipid metabolism. Cell Metab. 2020; 32:1052–62.
Article54. Mori Y, Ajay AK, Chang JH, Mou S, Zhao H, Kishi S, et al. KIM-1 mediates fatty acid uptake by renal tubular cells to promote progressive diabetic kidney disease. Cell Metab. 2021; 33:1042–61.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of Free Fatty Acid on Insulin Secretion in Cultured Rat Pancreatic Islets
- AMPK Activator AICAR Inhibits Hepatic Gluconeogenesis and Fatty Acid Oxidation
- Increase in Fatty Acid Oxidation by AICAR: the Role of p38 MAPK
- Inhibition of Sarcoplasmic Reticulum Ca2+ Uptake by Pyruvate and Fatty Acid in H9c2 Cardiomyocytes: Implications for Diabetic Cardiomyopathy
- Effects of Lovastatin on Free Fatty Acid Oxidation in Cultured L6 Rat Skeletal Muscle Cells
