Specific kinesin and dynein molecules participate in the unconventional protein secretion of transmembrane proteins
- Affiliations
-
- 1Department of Pharmacology, Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, Seoul 03722, Korea
- 2Severance Biomedical Science Institute, Yonsei University College of Medicine, Seoul 03722, Korea
- KMID: 2559136
- DOI: http://doi.org/10.4196/kjpp.2024.28.5.435
Abstract
- Secretory proteins, including plasma membrane proteins, are generally known to be transported to the plasma membrane through the endoplasmic reticulum-to-Golgi pathway. However, recent studies have revealed that several plasma membrane proteins and cytosolic proteins lacking a signal peptide are released via an unconventional protein secretion (UcPS) route, bypassing the Golgi during their journey to the cell surface. For instance, transmembrane proteins such as the misfolded cystic fibrosis transmembrane conductance regulator (CFTR) protein and the Spike protein of coronaviruses have been observed to reach the cell surface through a UcPS pathway under cell stress conditions. Nevertheless, the precise mechanisms of the UcPS pathway, particularly the molecular machineries involving cytosolic motor proteins, remain largely unknown. In this study, we identified specific kinesins, namely KIF1A and KIF5A, along with cytoplasmic dynein, as critical players in the unconventional trafficking of CFTR and the SARS-CoV-2 Spike protein. Gene silencing results demonstrated that knockdown of KIF1A, KIF5A, and the KIF-associated adaptor protein SKIP, FYCO1 significantly reduced the UcPS of △F508-CFTR. Moreover, gene silencing of these motor proteins impeded the UcPS of the SARS-CoV-2 Spike protein. However, the same gene silencing did not affect the conventional Golgimediated cell surface trafficking of wild-type CFTR and Spike protein. These findings suggest that specific motor proteins, distinct from those involved in conventional trafficking, are implicated in the stress-induced UcPS of transmembrane proteins.
Keyword
Figure
-

Fig. 1 KIF1A and KIF5A are involved in the unconventional trafficking of ∆F508-CFTR but not in the conventional trafficking of WT-CFTR. Effects of KIF1A and KIF5A gene silencing on the unconventional trafficking of ∆F508-CFTR (A) and conventional trafficking of WT-CFTR (C). Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (50 nM each, 48 h) together with plasmids encoding ∆F508/WT-CFTR (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. Surface-proteins versus cell lysates were quantified respectively (B, D). (E) Efficacy of each target gene knockdown. Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. b, band B (core-glycosylated) CFTR; c, band C (complex-glycosylated) CFTR; CFTR, cystic fibrosis transmembrane conductance regulator; WT, wild-type; ER, endoplasmic reticulum; ns, not significant. **p < 0.01.

Fig. 2 KIF13A and KIF13B are not involved in both unconventional and conventional trafficking of ∆F508-CFTR. Effects of KIF13A and KIF13B gene silencing on the unconventional trafficking of ∆F508-CFTR (A) and the conventional trafficking of WT-CFTR (C). Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (50 nM each, 48 h) together with plasmids encoding ∆F508/WT-CFTR (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. Surface-proteins versus cell lysates were quantified respectively (B, D). (E) Efficacy of each target gene knockdown. Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. b, band B (core-glycosylated) CFTR; c, band C (complex-glycosylated) CFTR; CFTR, cystic fibrosis transmembrane conductance regulator; WT, wild-type; ER, endoplasmic reticulum; ns, not significant. **p < 0.01.
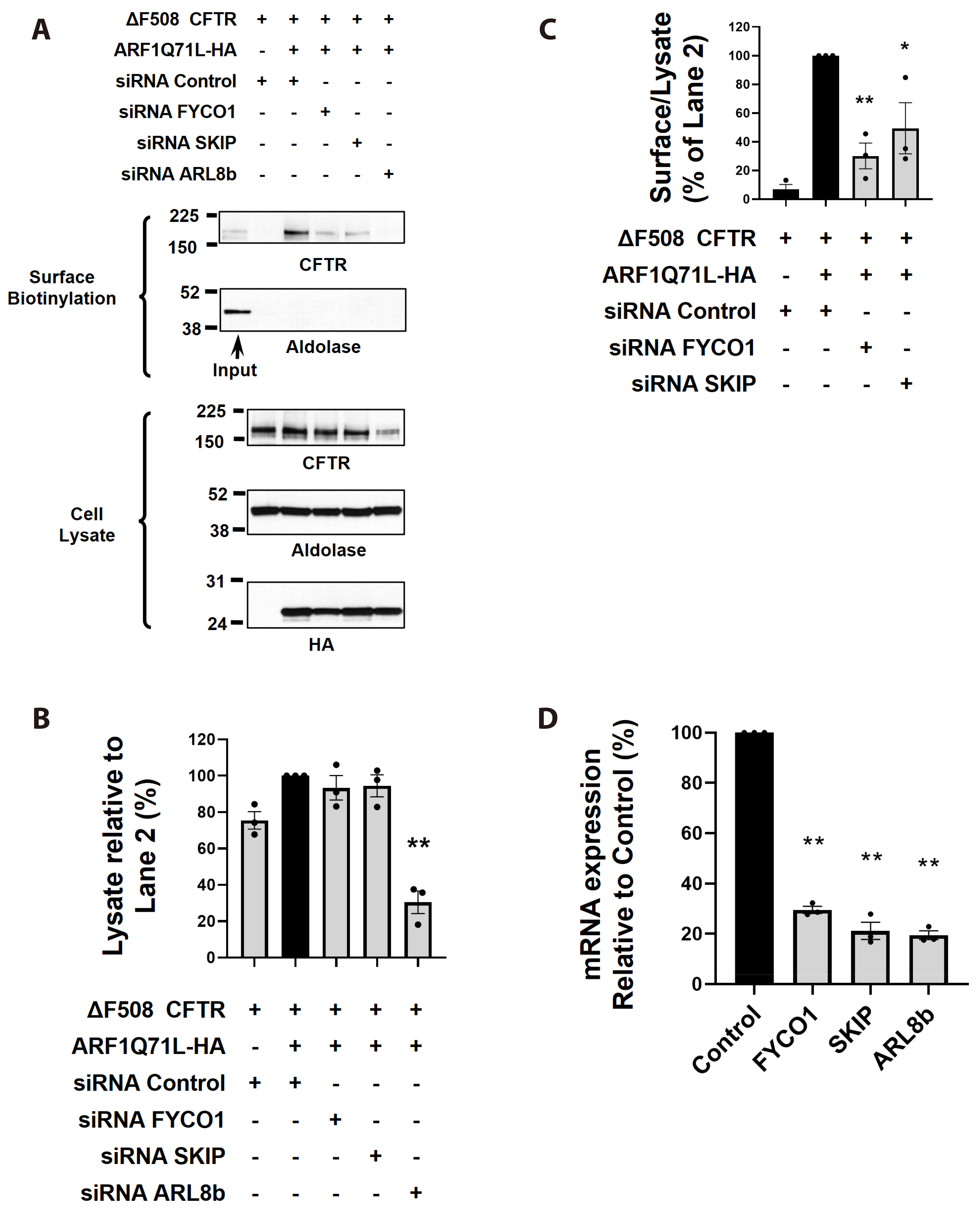
Fig. 3 FYCO1 and SKIP, acting as adaptor proteins, are involved in the unconventional trafficking of ∆F508-CFTR. Effects of SKIP, ARL8b, and FYCO1 gene silencing on the unconventional trafficking of ∆F508-CFTR. (A) Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (100 nM each, 48 h) together with plasmids encoding ∆F508-CFTR (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. (B) ∆F508-CFTR in lysate were quantified respectively. (C) Surface-proteins versus cell lysates were quantified respectively. (D) Efficacy of each target genes knockdown. Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. CFTR, cystic fibrosis transmembrane conductance regulator; ER, endoplasmic reticulum. *p < 0.05, **p < 0.01.

Fig. 4 Dynein is involved in the unconventional trafficking of ∆F508-CFTR but not in the conventional trafficking of WT-CFTR. Effects of each dynein subunit; DHC1, DIC1 and DLIC1 gene silencing on the unconventional trafficking of ∆F508-CFTR (A) and conventional trafficking of WT-CFTR (C). Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (100 nM each, 48 h) together with plasmids encoding ∆F508/WT-CFTR (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. Surface-proteins versus cell lysates were quantified respectively (B, D). (E) Efficacy of each target gene knockdown. Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. b, band B (core-glycosylated) CFTR; c, band C (complex-glycosylated) CFTR; CFTR, cystic fibrosis transmembrane conductance regulator; WT, wild-type; ER, endoplasmic reticulum; ns, not significant. **p < 0.01.

Fig. 5 Cell surface expression of ∆F508-CFTR is suppressed by knockdown of specific motor proteins. Immunofluorescent analysis was conducted on ΔF508-CFTR in cells subjected to siRNA treatment targeting KIF1A, KIF5A, and DHC1. Hela cells underwent sequential transfections with the indicated siRNAs (48 h) and plasmids (24 h), followed by immunostaining using anti-CFTR (green, Alexa Fluor 488) and anti-HA (red, Alexa Fluor 568) antibodies. The anti-HA immunofluorescence signal indicates the expression level of ARF1-Q71L. (A) ΔF508-CFTR was predominantly observed in perinuclear regions. (B) Co-transfection with ARF1Q71L in Hela cells resulted in an overall intracellular distribution and cell-surface expression of ΔF508-CFTR. Arrows represent the cell-surface rescue of ΔF508-CFTR by ARF1-Q71L. (C) Treatment with siRNAs against KIF1A (siKIF1A), (D) treatment with siRNAs against KIF5A (siKIF5A), and (E) treatment with siRNAs against DHC1 (siDHC1) abolished the ARF1Q71L-induced cell surface expression of the mutant CFTR. Three independent experiments yielded consistent results. Scale bar: 10 µm. CFTR, cystic fibrosis transmembrane conductance regulator.
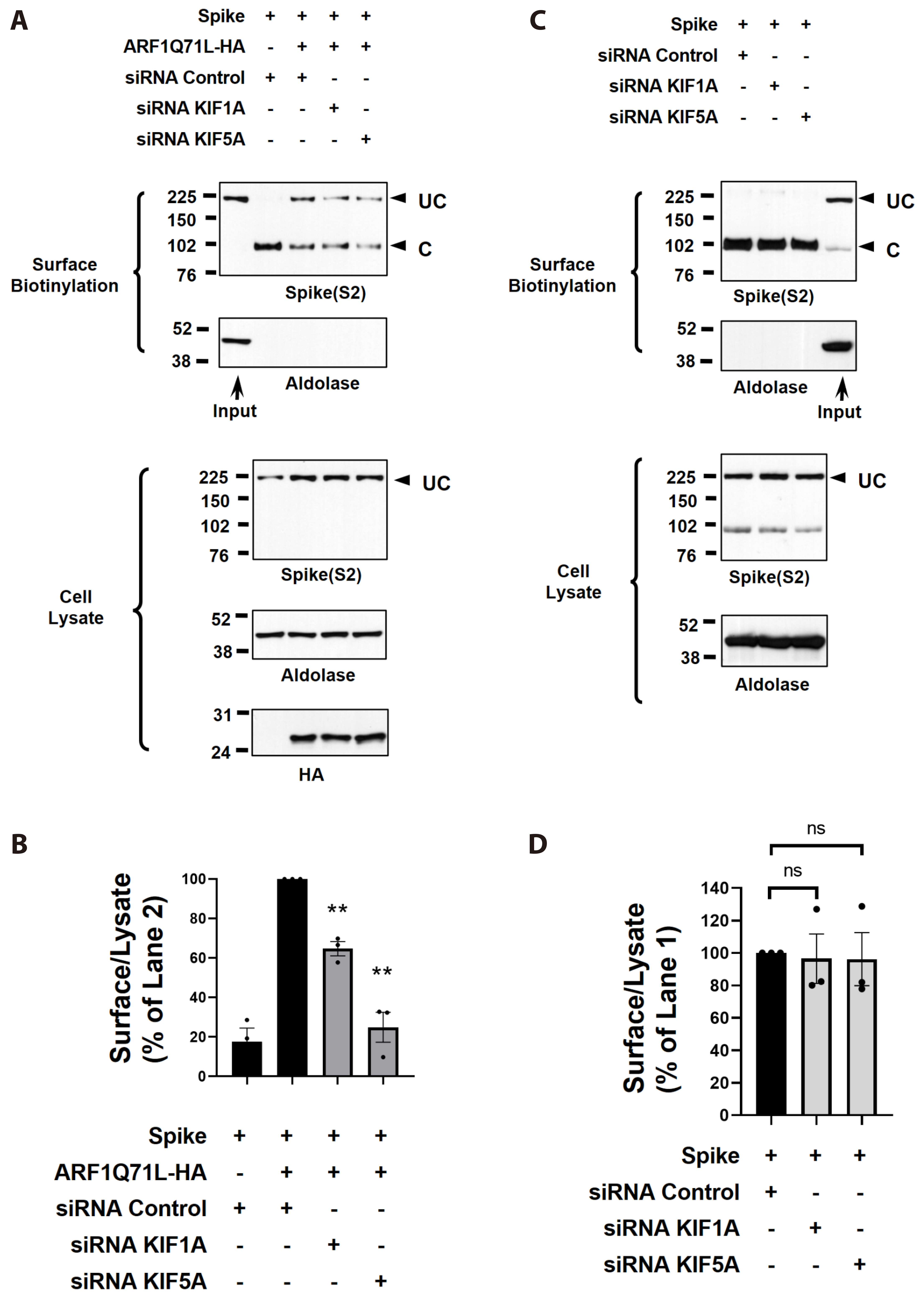
Fig. 6 KIF1A and KIF5A are involved in the unconventional trafficking of S protein but not in the conventional trafficking of S protein. Effects of KIF1A and KIF5A gene silencing on the unconventional trafficking of S protein (A) and conventional trafficking of S protein (C). Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (50 nM each, 48 h) together with plasmids encoding S protein (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. Surface-proteins versus cell lysates were quantified respectively (B, D). Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. UC, Golgi-bypassed uncleaved form of S protein; C, Golgi-dependent cleaved form of S protein; ER, endoplasmic reticulum; ns, not significant. **p < 0.01.

Fig. 7 Dynein is involved in the unconventional trafficking of S protein but not in the conventional trafficking of S protein. Effects of each dynein subunit; DHC1, DIC1, and DLIC1 gene silencing on the unconventional trafficking of S protein (A) and conventional trafficking of S protein (C). Cell surface biotinylation assay was conducted using HEK293 cells transfected with control (scrambled) siRNA or target siRNAs (100 nM each, 48 h) together with plasmids encoding S protein (24 h). ER-to-Golgi transport was inhibited by ARF1Q71L-HA overexpression in some cells. Surface-proteins versus cell lysates were quantified respectively (B, D). Bar graph data are presented as the mean ± SEM. Data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparison test. UC, Golgi-bypassed uncleaved form of S protein; C, Golgi-dependent cleaved form of S protein; ER, endoplasmic reticulum; ns, not significant. **p < 0.01.
Reference
-
1. Malhotra V. 2013; Unconventional protein secretion: an evolving mechanism. EMBO J. 32:1660–1664. DOI: 10.1038/emboj.2013.104. PMID: 23665917. PMCID: PMC3680731.2. Rabouille C. 2017; Pathways of unconventional protein secretion. Trends Cell Biol. 27:230–240. DOI: 10.1016/j.tcb.2016.11.007. PMID: 27989656.3. Gee HY, Noh SH, Tang BL, Kim KH, Lee MG. 2011; Rescue of ΔF508-CFTR trafficking via a GRASP-dependent unconventional secretion pathway. Cell. 146:746–760. DOI: 10.1016/j.cell.2011.07.021. PMID: 21884936.4. Park H, Seo SK, Sim JR, Hwang SJ, Kim YJ, Shin DH, Jang DG, Noh SH, Park PG, Ko SH, Shin MH, Choi JY, Ito Y, Kang CM, Lee JM, Lee MG. 2022; TMED3 complex mediates ER stress-associated secretion of CFTR, pendrin, and SARS-CoV-2 spike. Adv Sci (Weinh). 9:e2105320. DOI: 10.1002/advs.202105320. PMID: 35748162. PMCID: PMC9350134.5. O'Sullivan BP, Freedman SD. 2009; Cystic fibrosis. Lancet. 373:1891–1904. DOI: 10.1016/S0140-6736(09)60327-5. PMID: 19403164.6. Park H, Shin DH, Sim JR, Aum S, Lee MG. 2020; IRE1α kinase-mediated unconventional protein secretion rescues misfolded CFTR and pendrin. Sci Adv. 6:eaax9914. DOI: 10.1126/sciadv.aax9914. PMID: 32128399. PMCID: PMC7030921.7. Noh SH, Gee HY, Kim Y, Piao H, Kim J, Kang CM, Lee G, Mook-Jung I, Lee Y, Cho JW, Lee MG. 2018; Specific autophagy and ESCRT components participate in the unconventional secretion of CFTR. Autophagy. 14:1761–1778. DOI: 10.1080/15548627.2018.1489479. PMID: 29969945. PMCID: PMC6135621.8. Abraham Z, Hawley E, Hayosh D, Webster-Wood VA, Akkus O. 2018; Kinesin and dynein mechanics: measurement methods and research applications. J Biomech Eng. 140:0208051–02080511. DOI: 10.1115/1.4037886. PMID: 28901373. PMCID: PMC5816248.9. Hirokawa N, Noda Y, Tanaka Y, Niwa S. 2009; Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Biol. 10:682–696. DOI: 10.1038/nrm2774. PMID: 19773780.10. Roberts AJ. 2018; Emerging mechanisms of dynein transport in the cytoplasm versus the cilium. Biochem Soc Trans. 46:967–982. DOI: 10.1042/BST20170568. PMID: 30065109. PMCID: PMC6103457.11. Zhu C, Zhao J, Bibikova M, Leverson JD, Bossy-Wetzel E, Fan JB, Abraham RT, Jiang W. 2005; Functional analysis of human microtubule-based motor proteins, the kinesins and dyneins, in mitosis/cytokinesis using RNA interference. Mol Biol Cell. 16:3187–3199. DOI: 10.1091/mbc.e05-02-0167. PMID: 15843429. PMCID: PMC1165403.12. Urnavicius L, Zhang K, Diamant AG, Motz C, Schlager MA, Yu M, Patel NA, Robinson CV, Carter AP. 2015; The structure of the dynactin complex and its interaction with dynein. Science. 347:1441–1446. DOI: 10.1126/science.aaa4080. PMID: 25814576. PMCID: PMC4413427.13. Guardia CM, Farías GG, Jia R, Pu J, Bonifacino JS. 2016; BORC functions upstream of kinesins 1 and 3 to coordinate regional movement of lysosomes along different microtubule tracks. Cell Rep. 17:1950–1961. DOI: 10.1016/j.celrep.2016.10.062. PMID: 27851960. PMCID: PMC5136296.14. Delevoye C, Hurbain I, Tenza D, Sibarita JB, Uzan-Gafsou S, Ohno H, Geerts WJ, Verkleij AJ, Salamero J, Marks MS, Raposo G. 2009; AP-1 and KIF13A coordinate endosomal sorting and positioning during melanosome biogenesis. J Cell Biol. 187:247–264. DOI: 10.1083/jcb.200907122. PMID: 19841138. PMCID: PMC2768840.15. Etoh K, Fukuda M. 2019; Rab10 regulates tubular endosome formation through KIF13A and KIF13B motors. J Cell Sci. 132:jcs226977. DOI: 10.1242/jcs.226977. PMID: 30700496.16. Tong Y, Tempel W, Wang H, Yamada K, Shen L, Senisterra GA, MacKenzie F, Chishti AH, Park HW. 2010; Phosphorylation-independent dual-site binding of the FHA domain of KIF13 mediates phosphoinositide transport via centaurin alpha1. Proc Natl Acad Sci U S A. 107:20346–20351. DOI: 10.1073/pnas.1009008107. PMID: 21057110. PMCID: PMC2996657.17. Cross JA, Dodding MP. 2019; Motor-cargo adaptors at the organelle-cytoskeleton interface. Curr Opin Cell Biol. 59:16–23. DOI: 10.1016/j.ceb.2019.02.010. PMID: 30952037.18. Roney JC, Cheng XT, Sheng ZH. 2022; Neuronal endolysosomal transport and lysosomal functionality in maintaining axonostasis. J Cell Biol. 221:e202111077. DOI: 10.1083/jcb.202111077. PMID: 35142819. PMCID: PMC8932522.19. Sanger A, Yip YY, Randall TS, Pernigo S, Steiner RA, Dodding MP. 2017; SKIP controls lysosome positioning using a composite kinesin-1 heavy and light chain-binding domain. J Cell Sci. 130:1637–1651. DOI: 10.1242/jcs.198267. PMID: 28302907. PMCID: PMC5450233.20. Farías GG, Guardia CM, De Pace R, Britt DJ, Bonifacino JS. 2017; BORC/kinesin-1 ensemble drives polarized transport of lysosomes into the axon. Proc Natl Acad Sci U S A. 114:E2955–E2964. DOI: 10.1073/pnas.1616363114. PMID: 28320970. PMCID: PMC5389300.21. Khatter D, Sindhwani A, Sharma M. 2015; Arf-like GTPase Arl8: moving from the periphery to the center of lysosomal biology. Cell Logist. 5:e1086501. DOI: 10.1080/21592799.2015.1086501. PMID: 27057420. PMCID: PMC4820812.22. Pankiv S, Alemu EA, Brech A, Bruun JA, Lamark T, Overvatn A, Bjørkøy G, Johansen T. 2010; FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end-directed vesicle transport. J Cell Biol. 188:253–269. DOI: 10.1083/jcb.200907015. PMID: 20100911. PMCID: PMC2812517.23. Matsuzaki F, Shirane M, Matsumoto M, Nakayama KI. 2011; Protrudin serves as an adaptor molecule that connects KIF5 and its cargoes in vesicular transport during process formation. Mol Biol Cell. 22:4602–4620. DOI: 10.1091/mbc.e11-01-0068. PMID: 21976701. PMCID: PMC3226478.24. Toropova K, Zalyte R, Mukhopadhyay AG, Mladenov M, Carter AP, Roberts AJ. 2019; Structure of the dynein-2 complex and its assembly with intraflagellar transport trains. Nat Struct Mol Biol. 26:823–829. DOI: 10.1038/s41594-019-0286-y. PMID: 31451806. PMCID: PMC6774794.25. Reck-Peterson SL, Redwine WB, Vale RD, Carter AP. 2018; The cytoplasmic dynein transport machinery and its many cargoes. Nat Rev Mol Cell Biol. 19:382–398. Erratum in: Nat Rev Mol Cell Biol. 2018;19:479. DOI: 10.1038/s41580-018-0004-3. PMID: 29662141. PMCID: PMC6457270.26. Kim J, Noh SH, Piao H, Kim DH, Kim K, Cha JS, Chung WY, Cho HS, Kim JY, Lee MG. 2016; Monomerization and ER relocalization of GRASP is a requisite for unconventional secretion of CFTR. Traffic. 17:733–753. DOI: 10.1111/tra.12403. PMID: 27062250.27. Piao H, Kim J, Noh SH, Kweon HS, Kim JY, Lee MG. 2017; Sec16A is critical for both conventional and unconventional secretion of CFTR. Sci Rep. 7:39887. DOI: 10.1038/srep39887. PMID: 28067262. PMCID: PMC5220342.28. Watanabe Y, Allen JD, Wrapp D, McLellan JS, Crispin M. 2020; Site-specific glycan analysis of the SARS-CoV-2 spike. Science. 369:330–333. DOI: 10.1126/science.abb9983. PMID: 32366695. PMCID: PMC7199903.29. Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, Guo L, Guo R, Chen T, Hu J, Xiang Z, Mu Z, Chen X, Chen J, Hu K, Jin Q, Wang J, Qian Z. 2020; Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. 11:1620. Erratum in: Nat Commun. 2021;12:2144. DOI: 10.1038/s41467-021-22614-1. PMID: 33795662. PMCID: PMC8016153.30. Maday S, Twelvetrees AE, Moughamian AJ, Holzbaur EL. 2014; Axonal transport: cargo-specific mechanisms of motility and regulation. Neuron. 84:292–309. DOI: 10.1016/j.neuron.2014.10.019. PMID: 25374356. PMCID: PMC4269290.31. Burgo A, Proux-Gillardeaux V, Sotirakis E, Bun P, Casano A, Verraes A, Liem RK, Formstecher E, Coppey-Moisan M, Galli T. 2012; A molecular network for the transport of the TI-VAMP/VAMP7 vesicles from cell center to periphery. Dev Cell. 23:166–80. DOI: 10.1016/j.devcel.2012.04.019. PMID: 22705394.32. Kamata H, Tsukasaki Y, Sakai T, Ikebe R, Wang J, Jeffers A, Boren J, Owens S, Suzuki T, Higashihara M, Idell S, Tucker TA, Ikebe M. 2017; KIF5A transports collagen vesicles of myofibroblasts during pleural fibrosis. Sci Rep. 7:4556. DOI: 10.1038/s41598-017-04437-7. PMID: 28676645. PMCID: PMC5496869.33. Nakajima K, Yin X, Takei Y, Seog DH, Homma N, Hirokawa N. 2012; Molecular motor KIF5A is essential for GABA(A) receptor transport, and KIF5A deletion causes epilepsy. Neuron. 76:945–961. DOI: 10.1016/j.neuron.2012.10.012. PMID: 23217743.34. Ghosh S, Dellibovi-Ragheb TA, Kerviel A, Pak E, Qiu Q, Fisher M, Takvorian PM, Bleck C, Hsu VW, Fehr AR, Perlman S, Achar SR, Straus MR, Whittaker GR, de Haan CAM, Kehrl J, Altan-Bonnet G, Altan-Bonnet N. 2020; β-coronaviruses use lysosomes for egress instead of the biosynthetic secretory pathway. Cell. 183:1520–1535.e14. DOI: 10.1016/j.cell.2020.10.039. PMID: 33157038. PMCID: PMC7590812.35. D'Souza AI, Grover R, Monzon GA, Santen L, Diez S. 2023; Vesicles driven by dynein and kinesin exhibit directional reversals without regulators. Nat Commun. 14:7532. Erratum in: Nat Commun. 2024;15:736. DOI: 10.1038/s41467-023-42605-8. PMID: 37985763. PMCID: PMC10662051.36. Watanabe TM, Higuchi H. 2007; Stepwise movements in vesicle transport of HER2 by motor proteins in living cells. Biophys J. 92:4109–4120. DOI: 10.1529/biophysj.106.094649. PMID: 17369416. PMCID: PMC1868978.37. Sleigh JN, Rossor AM, Fellows AD, Tosolini AP, Schiavo G. 2019; Axonal transport and neurological disease. Nat Rev Neurol. 15:691–703. DOI: 10.1038/s41582-019-0257-2. PMID: 31558780.38. Gönczy P, Pichler S, Kirkham M, Hyman AA. 1999; Cytoplasmic dynein is required for distinct aspects of MTOC positioning, including centrosome separation, in the one cell stage Caenorhabditis elegans embryo. J Cell Biol. 147:135–150. DOI: 10.1083/jcb.147.1.135. PMID: 10508861. PMCID: PMC2164971.39. Fourriere L, Jimenez AJ, Perez F, Boncompain G. 2020; The role of microtubules in secretory protein transport. J Cell Sci. 133:jcs237016. DOI: 10.1242/jcs.237016. PMID: 31996399.40. Bruns C, McCaffery JM, Curwin AJ, Duran JM, Malhotra V. 2011; Biogenesis of a novel compartment for autophagosome-mediated unconventional protein secretion. J Cell Biol. 195:979–992. DOI: 10.1083/jcb.201106098. PMID: 22144692. PMCID: PMC3241719.41. Woźniak MJ, Bola B, Brownhill K, Yang YC, Levakova V, Allan VJ. 2009; Role of kinesin-1 and cytoplasmic dynein in endoplasmic reticulum movement in VERO cells. J Cell Sci. 122:1979–1989. DOI: 10.1242/jcs.041962. PMID: 19454478. PMCID: PMC2723153.42. Lu M, van Tartwijk FW, Lin JQ, Nijenhuis W, Parutto P, Fantham M, Christensen CN, Avezov E, Holt CE, Tunnacliffe A, Holcman D, Kapitein L, Schierle GSK, Kaminski CF. 2020; The structure and global distribution of the endoplasmic reticulum network are actively regulated by lysosomes. Sci Adv. 6:eabc7209. DOI: 10.1126/sciadv.abc7209. PMID: 33328230. PMCID: PMC7744115.43. Pu J, Schindler C, Jia R, Jarnik M, Backlund P, Bonifacino JS. 2015; BORC, a multisubunit complex that regulates lysosome positioning. Dev Cell. 33:176–188. DOI: 10.1016/j.devcel.2015.02.011. PMID: 25898167. PMCID: PMC4788105.44. Pu J, Guardia CM, Keren-Kaplan T, Bonifacino JS. 2016; Mechanisms and functions of lysosome positioning. J Cell Sci. 129:4329–4339. DOI: 10.1242/jcs.196287. PMID: 27799357. PMCID: PMC5201012.45. Jung J, Kim J, Roh SH, Jun I, Sampson RD, Gee HY, Choi JY, Lee MG. 2016; The HSP70 co-chaperone DNAJC14 targets misfolded pendrin for unconventional protein secretion. Nat Commun. 7:11386. DOI: 10.1038/ncomms11386. PMID: 27109633. PMCID: PMC4848490.46. Monteleone M, Stow JL, Schroder K. 2015; Mechanisms of unconventional secretion of IL-1 family cytokines. Cytokine. 74:213–218. DOI: 10.1016/j.cyto.2015.03.022. PMID: 25922276.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Sorting Nexin 17 Interacts Directly with Kinesin Superfamily KIF1B beta Protein
- Molecular Motor Proteins of the Kinesin Superfamily Proteins (KIFs): Structure, Cargo and Disease
- Kinesin Spindle Protein Inhibition in Translational Research
- Corrigendum: Figure Correction. Primary Cilia as a Signaling Platform for Control of Energy Metabolism
- Transmembrane Adaptor Proteins Positively Regulating the Activation of Lymphocytes
