Umami taste receptor suppresses cancer cachexia by regulating skeletal muscle atrophy in vivo and in vitro
- Affiliations
-
- 1Department of Nutritional Science and Food Management, Ewha Womans University, Seoul 03760, Korea
- 2Interdisciplinary Program in Bioengineering, Seoul National University, Seoul 08826, Korea
- 3Graduate Program in System Health Science and Engineering, Ewha Womans University, Seoul 03760, Korea
- KMID: 2558485
- DOI: http://doi.org/10.4162/nrp.2024.18.4.451
Abstract
- BACKGROUND/OBJECTIVES
The umami taste receptor (TAS1R1/TAS1R3) is endogenously expressed in skeletal muscle and is involved in myogenesis; however, there is a lack of evidence about whether the expression of the umami taste receptor is involved in muscular diseases. This study aimed to elucidate the effects of the umami taste receptor and its mechanism on muscle wasting in cancer cachexia using in vivo and in vitro models.
MATERIALS/METHODS
The Lewis lung carcinoma-induced cancer cachexia model was used in vivo and in vitro, and the expressions of umami taste receptor and muscle atrophy-related markers, muscle atrophy F-box protein, and muscle RING-finger protein-1 were analyzed.
RESULTS
Results showed that TAS1R1 was significantly downregulated in vivo and in vitro under the muscle wasting condition. Moreover, overexpression of TAS1R1 in vitro in the human primary cell model protected the cells from muscle atrophy, and knockdown of TAS1R1 using siRNA exacerbated muscle atrophy.
CONCLUSION
Taken together, the umami taste receptor exerts protective effects on muscle-wasting conditions by restoring dysregulated muscle atrophy in cancer cachexia. In conclusion, this result provided evidence that the umami taste receptor exerts a therapeutic anti-cancer cachexia effect by restoring muscle atrophy.
Keyword
Figure
-
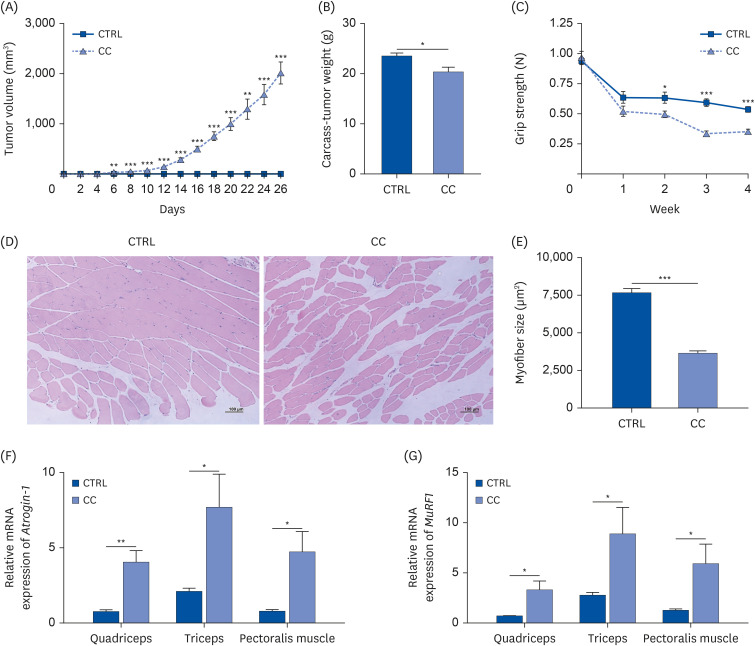
Fig. 1 Muscle atrophic features in LLC-induced cancer cachexia mouse model. (A) Tumor volumes, (B) carcass weights after tumor removal (carcass-tumor weight), and (C) grip strength were measured (n = 6/group). (D) The representative quadriceps tissues stained with H&E were shown (magnification 100×, scale bar 100 μm), and (E) their myofiber size was quantified (n = 3/group). mRNA expressions of muscle atrophy-related genes (F) Atrogin-1 and (G) MuRF1 in quadriceps, triceps, and pectoralis muscle were determined by RT-qPCR (n = 6/group). All data were shown as mean ± SEM and analyzed by an unpaired 2-tailed Student’s t-test.CTRL, control; CC, LLC-induced cancer cachexia; H&E, hematoxylin and eosin. P < 0.05 was considered statistically significant (*P < 0.05, **P < 0.01, ***P < 0.001).
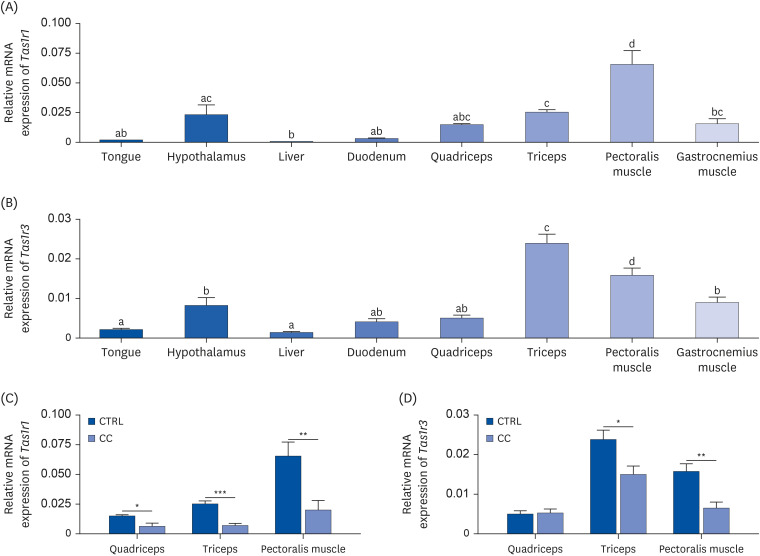
Fig. 2 Umami taste receptor is expressed in different types of skeletal muscle and is downregulated in cachexia condition in an in vivo mouse model. The mRNA expressions level of (A) Tas1r1 and (B) Tas1r3 were analyzed in mouse tongue, hypothalamus, liver, duodenum, quadriceps, triceps, pectoralis muscle, and gastrocnemius muscles of the CTRL group (n = 6/group). The mRNA expressions of (C) Tas1r1 and (D) Tas1r3 in quadriceps, triceps, and pectoralis muscles were analyzed. β-actin was used as a loading control (n = 6/group). Data were analyzed by one-way analysis of variance with a Newman-Keuls’s post hoc test. Different letters indicate statistical significance (P < 0.05), and an unpaired 2-tailed Student’s t-test was performed to compare data between 2 groups. All bars represent the mean ± SEM.CTRL, control; CC, LLC-induced cancer cachexia.P < 0.05 was considered statistically significant (*P < 0.05, **P < 0.01, ***P < 0.001).
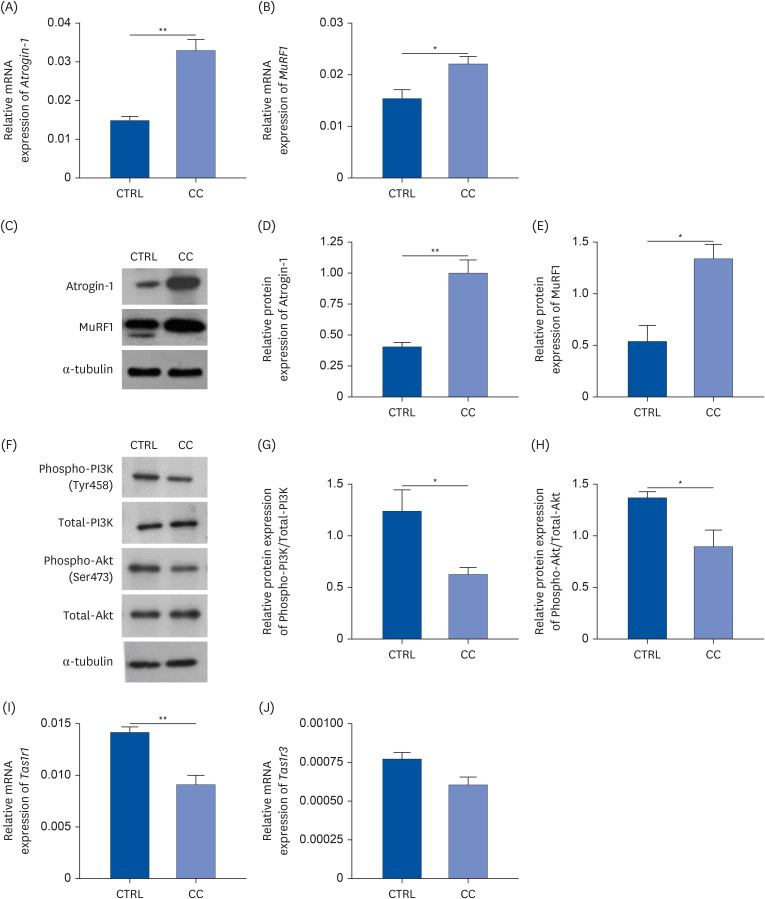
Fig. 3 Muscle atrophy markers are upregulated and the umami taste receptor is downregulated in cachexia conditions in an in vitro mouse model. The mRNA levels of (A) Atrogin-1 and (B) MuRF1 were analyzed by RT-qPCR. The protein levels of (C-E) Atrogin-1, MuRF1, and (F-H) PI3K/Akt pathway in C2C12 cells were analyzed by Western blotting. (C, F) Representative blots were shown. (D, E, G, H) The relative band intensities were calculated after normalization to α-tubulin expression. The mRNA levels of (I) Tas1r1 and (J) Tas1r3 were measured by RT-qPCR. All values were shown as mean ± SEM (n = 3). Data were analyzed by an unpaired 2-tailed Student’s t-test.CTRL, control; CC, LLC-induced cancer cachexia; MuRF1, muscle RING-finger protein-1; RT-qPCR, reverse transcription-quantitative polymerase chain reaction; PI3K, phosphoinositide 3-kinase. P < 0.05 was considered statistically significant (*P < 0.05, **P < 0.01).
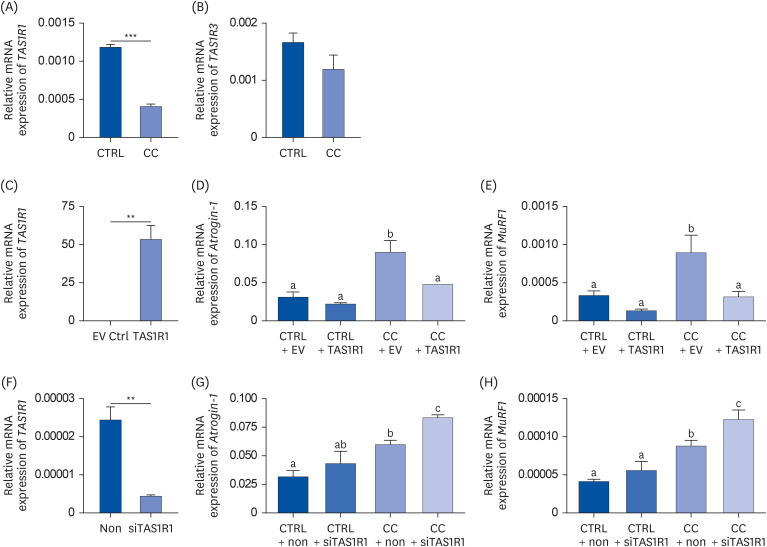
Fig. 4 Umami taste receptor is downregulated in cachexia conditions in an in vitro human model and suppresses atrophic markers. The mRNA levels of (A) TAS1R1 and (B) TAS1R3 were analyzed by RT-qPCR. (C-E) Fully differentiated HSkM cells were transfected with pCMV6 EV or pCMV6-Entry containing TAS1R1. The mRNA levels of (C) TAS1R1, (D) Atrogin-1, and (E) MuRF1 were analyzed by RT-qPCR. (F-H) Fully differentiated HSkM cells were transfected with control siRNA or TAS1R1 siRNA. The mRNA levels of (F) TAS1R1, (G) Atrogin-1, and (H) MuRF1 were measured by RT-qPCR. GAPDH was used as a loading control and all experiment results are shown as mean ± SEM (n = 3). Data were analyzed by one-way analysis of variance with a Newman-Keuls’s post hoc test. Different letters indicate statistical significance (P < 0.05), and an unpaired 2-tailed Student’s t-test was performed to compare data between 2 groups.CTRL, control; CC, LLC-induced cancer cachexia; EV Ctrl, empty vector control; CTRL + EV, control + empty vector; CTRL + TAS1R1, control + TAS1R1; CC + EV, LLC-induced cancer cachexia + empty vector; CC + TAS1R1, LLC-induced cancer cachexia + TAS1R1; non, non-targeting siRNA; CTRL + non, control + non-targeting siRNA; CTRL + siTAS1R1, control + TAS1R1 siRNA; CC + non, LLC-induced cancer cachexia + non-targeting siRNA; CC + siTAS1R1, LLC-induced cancer cachexia + TAS1R1 siRNA; MuRF1, muscle RING-finger protein-1; RT-qPCR, reverse transcription-quantitative polymerase chain reaction. P < 0.05 was considered statistically significant (**P < 0.01, ***P < 0.001).
Reference
-
1. Fearon K, Strasser F, Anker SD, Bosaeus I, Bruera E, Fainsinger RL, Jatoi A, Loprinzi C, MacDonald N, Mantovani G, et al. Definition and classification of cancer cachexia: an international consensus. Lancet Oncol. 2011; 12:489–495. PMID: 21296615.
Article2. Kasvis P, Vigano M, Vigano A. Health-related quality of life across cancer cachexia stages. Ann Palliat Med. 2019; 8:33–42. PMID: 30525763.3. Dodson S, Baracos VE, Jatoi A, Evans WJ, Cella D, Dalton JT, Steiner MS. Muscle wasting in cancer cachexia: clinical implications, diagnosis, and emerging treatment strategies. Annu Rev Med. 2011; 62:265–279. PMID: 20731602.
Article4. Balagopal P, Ljungqvist O, Nair KS. Skeletal muscle myosin heavy-chain synthesis rate in healthy humans. Am J Physiol. 1997; 272:E45–E50. PMID: 9038850.
Article5. Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, Poueymirou WT, Panaro FJ, Na E, Dharmarajan K, et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001; 294:1704–1708. PMID: 11679633.
Article6. Yuan L, Han J, Meng Q, Xi Q, Zhuang Q, Jiang Y, Han Y, Zhang B, Fang J, Wu G. Muscle-specific E3 ubiquitin ligases are involved in muscle atrophy of cancer cachexia: an in vitro and in vivo study. Oncol Rep. 2015; 33:2261–2268. PMID: 25760630.
Article7. Adams V, Gußen V, Zozulya S, Cruz A, Moriscot A, Linke A, Labeit S. Small-molecule chemical knockdown of MuRF1 in melanoma bearing mice attenuates tumor cachexia associated myopathy. Cells. 2020; 9:2272. PMID: 33050629.8. Moro T, Ebert SM, Adams CM, Rasmussen BB. Amino acid sensing in skeletal muscle. Trends Endocrinol Metab. 2016; 27:796–806. PMID: 27444066.
Article9. Wauson EM, Zaganjor E, Lee AY, Guerra ML, Ghosh AB, Bookout AL, Chambers CP, Jivan A, McGlynn K, Hutchison MR, et al. The G protein-coupled taste receptor T1R1/T1R3 regulates mTORC1 and autophagy. Mol Cell. 2012; 47:851–862. PMID: 22959271.
Article10. Wauson EM, Lorente-Rodríguez A, Cobb MH. Minireview: nutrient sensing by G protein-coupled receptors. Mol Endocrinol. 2013; 27:1188–1197. PMID: 23820899.
Article11. Shirakawa T, Toyono T, Inoue A, Matsubara T, Kawamoto T, Kokabu S. Factors regulating or regulated by myogenic regulatory factors in skeletal muscle stem cells. Cells. 2022; 11:1493. PMID: 35563799.
Article12. Kokabu S, Lowery JW, Toyono T, Seta Y, Hitomi S, Sato T, Enoki Y, Okubo M, Fukushima Y, Yoda T. Muscle regulatory factors regulate T1R3 taste receptor expression. Biochem Biophys Res Commun. 2015; 468:568–573. PMID: 26545778.13. Obikane Y, Toyono T, Kokabu S, Matsuyama K, Kataoka S, Nakatomi M, Hosokawa R, Seta Y. Myogenic differentiation 1 and transcription factor 12 activate the gene expression of mouse taste receptor type 1 member 1. J Oral Biosci. 2021; 63:420–428. PMID: 34492379.
Article14. Carlsson G, Gullberg B, Hafström L. Estimation of liver tumor volume using different formulas - an experimental study in rats. J Cancer Res Clin Oncol. 1983; 105:20–23. PMID: 6833336.
Article15. Ahn SR, An JH, Song HS, Park JW, Lee SH, Kim JH, Jang J, Park TH. Duplex bioelectronic tongue for sensing umami and sweet tastes based on human taste receptor nanovesicles. ACS Nano. 2016; 10:7287–7296. PMID: 27327579.
Article16. Dalesio NM, Barreto Ortiz SF, Pluznick JL, Berkowitz DE. Olfactory, taste, and photo sensory receptors in non-sensory organs: it just makes sense. Front Physiol. 2018; 9:1673. PMID: 30542293.
Article17. Li X, Staszewski L, Xu H, Durick K, Zoller M, Adler E. Human receptors for sweet and umami taste. Proc Natl Acad Sci U S A. 2002; 99:4692–4696. PMID: 11917125.18. Kokabu S, Lowery JW, Toyono T, Sato T, Yoda T. On the emerging role of the taste receptor type 1 (T1R) family of nutrient-sensors in the musculoskeletal system. Molecules. 2017; 22:469. PMID: 28294983.
Article19. Rask-Andersen M, Almén MS, Schiöth HB. Trends in the exploitation of novel drug targets. Nat Rev Drug Discov. 2011; 10:579–590. PMID: 21804595.
Article20. Xu H, Staszewski L, Tang H, Adler E, Zoller M, Li X. Different functional roles of T1R subunits in the heteromeric taste receptors. Proc Natl Acad Sci U S A. 2004; 101:14258–14263. PMID: 15353592.
Article21. Schiaffino S, Dyar KA, Ciciliot S, Blaauw B, Sandri M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013; 280:4294–4314. PMID: 23517348.
Article22. Rommel C, Bodine SC, Clarke BA, Rossman R, Nunez L, Stitt TN, Yancopoulos GD, Glass DJ. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat Cell Biol. 2001; 3:1009–1013. PMID: 11715022.23. Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A, Walsh K, Schiaffino S, Lecker SH, Goldberg AL. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell. 2004; 117:399–412. PMID: 15109499.
Article
