Somatodendritic organization of pacemaker activity in midbrain dopamine neurons
- Affiliations
-
- 1Department of Physiology, Sungkyunkwan University School of Medicine, Suwon 16419, Korea
- KMID: 2553528
- DOI: http://doi.org/10.4196/kjpp.2024.28.2.165
Abstract
- The slow and regular pacemaking activity of midbrain dopamine (DA) neurons requires proper spatial organization of the excitable elements between the soma and dendritic compartments, but the somatodendritic organization is not clear. Here, we show that the dynamic interaction between the soma and multiple proximal dendritic compartments (PDCs) generates the slow pacemaking activity in DA neurons. In multipolar DA neurons, spontaneous action potentials (sAPs) consistently originate from the axon-bearing dendrite. However, when the axon initial segment was disabled, sAPs emerge randomly from various primary PDCs, indicating that multiple PDCs drive pacemaking. Ca2+ measurements and local stimulation/perturbation experiments suggest that the soma serves as a stably-oscillating inertial compartment, while multiple PDCs exhibit stochastic fluctuations and high excitability. Despite the stochastic and excitable nature of PDCs, their activities are balanced by the large centrally-connected inertial soma, resulting in the slow synchronized pacemaking rhythm. Furthermore, our electrophysiological experiments indicate that the soma and PDCs, with distinct characteristics, play different roles in glutamate-induced burst-pause firing patterns. Excitable PDCs mediate excitatory burst responses to glutamate, while the large inertial soma determines inhibitory pause responses to glutamate. Therefore, we could conclude that this somatodendritic organization serves as a common foundation for both pacemaker activity and evoked firing patterns in midbrain DA neurons.
Keyword
Figure
-

Fig. 1 Axon removal reveals dendritic origin of sAPs in DA neurons. (A) Fluorescence images of a DA neuron in a midbrain slice and pacemaking and spontaneous firing diagram. The yellow rectangular area containing an axon is expanded into the middle images before and after axon removal at the red line. Right, spontaneous firing traces recorded in the cell-attached mode before and after axon removal. (B) Mean spontaneous firing frequencies before and after axon removal. (C) ISI variances before and after axon removal. (D) Ca2+ spikes were measured in two opposite proximal primary dendrites, P1 (ABD, blue circles) and P2 (nABD, red circles). (E) Before axon-cutting, Ca2+ spikes always developed immediately after sAPs from the ABD (P1, blue) and later at the nABD (P2, red). Black line represents AP appearance timing. AP propagation direction was marked with arrow in an insert. (F) Time-lags between APs and rising-points of Ca2+ spikes (mean time lag: P1 = 0.473 ± 0.046, P2 = 10.39 ± 1.04, n = 3). (G) Variances in the time-lags before and after axon removal. (H) Ca2+ spikes were measured in two opposite proximal primary dendrites (P1 and P2) after axon removal. The order of the two dendritic Ca2+ spikes varied. The case in which the Ca2+ spike occurred first at the nABD, immediately after sAP, is presented. (I) The order of Ca2+ spikes between two primary dendrites with the time-lags (mean time lag: P1 = 10.86 ± 2.69, P2 = 6.85 ± 2.19, n = 3). (J) Incidence of AP initiation in the ABD (P1) and nABD (P2) changed dramatically after axon removal (from P1 = 100% and P2 = 0% to P1 = 55.56%, and P2 = 44.44%, n = 3). Values are presented as mean ± SE. DA, dopamine; ISI, interspike interval; ABD, axon-bearing dendrite; nABD, non-axon-bearing dendrite; AP, action potential; sAPs, spontaneous action potentials; AIS, axon initial segment. **p < 0.001.
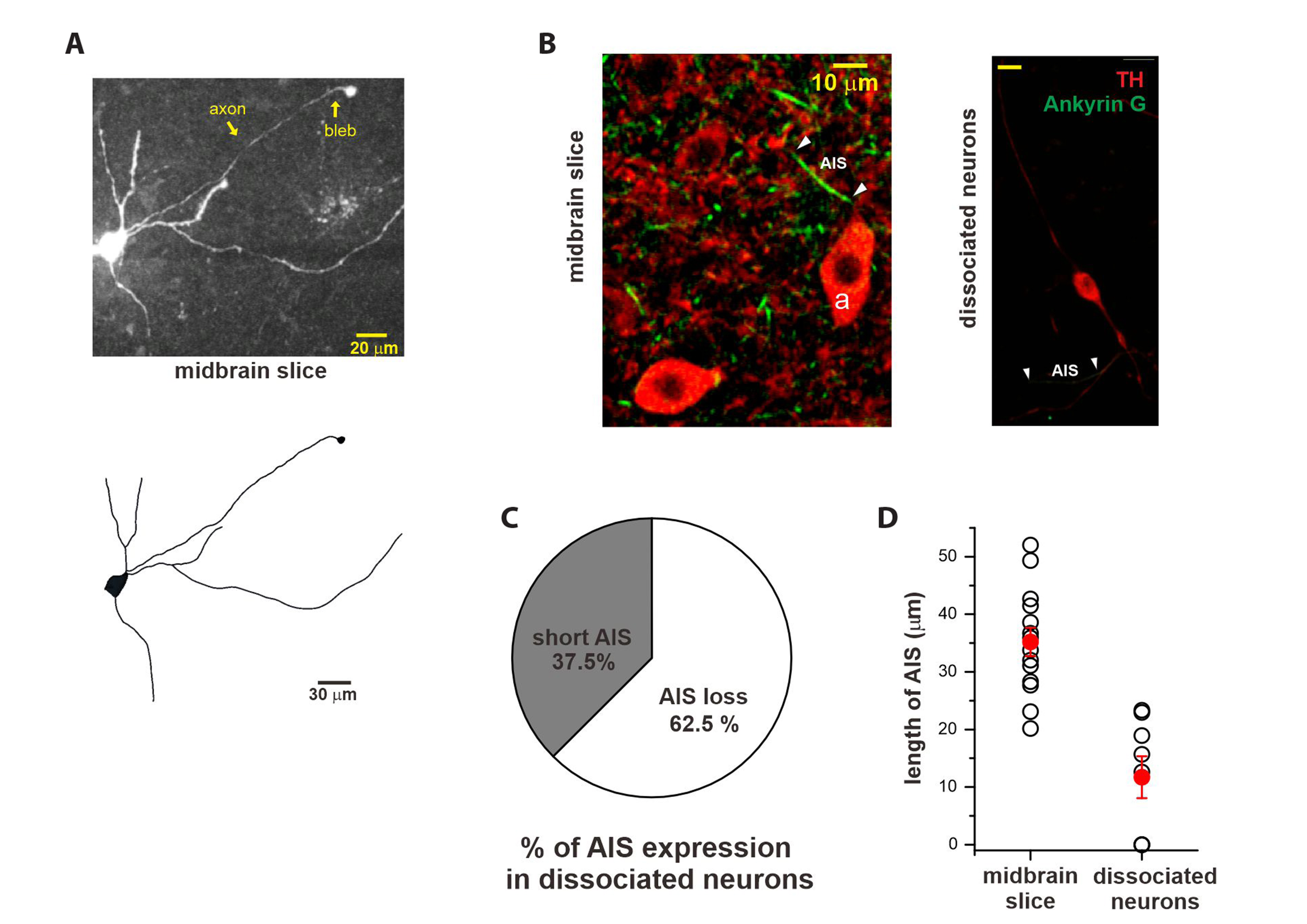
Fig. 2 Comparison of the AIS between midbrain slices and dissociated DA neurons. (A) AISs were identified in the DA neurons in the midbrain slices using two-photon confocal imaging and a terminal retraction ball formed during slice preparation. Bottom, the simplified structure of a DA neuron that includes a thin axon and multiple dendrites is reconstructed from the upper fluorescence image. (B) Immunocytochemistry of TH (red; Alexa 647) and ankyrin G (green; Qdot 488) in a SNc brain slice and dissociated DA neurons. The AISs of one DA neuron marked as ʻaʼ in the slice and a dissociated neuron are marked by white triangles. (C) Significant loss of AIS in dissociated DA neurons. AIS loss in 62.5% (5/8 cells) and decreased AIS in 37.5% (3/8 cells). (D) Average AIS length in the dissociated DA neurons decreased significantly after dissociation procedures, as shown by comparing them with DA neurons in the slices (AIS lengths of DA neurons in slices, 35.20 ± 2.26 µm, n = 15; in dissociated DA neurons, 11.70 ± 3.64 µm, n = 8). AIS, axon initial segment; DA, dopamine; TH, tyrosine hydroxylase; SNc, substantia nigra pars compacta.

Fig. 3 Variable initiation of sAPs from multiple PDCs in the primary dendrites in acutely dissociated, AIS-disabled DA neurons. (A) The red fluorescence image with differential interference contrast image of an acutely dissociated DA neuron loaded with Fluo 4-AM and Ca2+ measuring sites (S, soma; P, proximal dendrite; and D, distal dendrite). (B) After sAP, the Ca2+ spike started immediately from the PDC (red) and then propagated into the soma (black) and distal dendritic region (blue). Normalized traces expanded from the inset. (C) The Ca2+ spike started from the soma and propagated directionally to the proximal and distal dendritic regions. Normalized traces expanded from the inset. (D) Distribution of the time-lags of Ca2+ spike onsets between the dendritic regions and the soma. Black circles indicate Ca2+ propagation from the soma to the dendrite. Red circles indicate Ca2+ propagation from the proximal dendrite to the soma. Gray closed circles indicate time-lags in the distal dendritic region. Right, time lags of Ca2+ onset are represented by a bar graph (P→S: –6.06 ± 1.66 ms, 13/24, S→P: 6.75 ± 2.33 ms, 11/24). (E) Initiation order of Ca2+ spikes among three different parts, including two opposite proximal primary dendrites. Two individual PDCs, P1 (red closed circles) and P2 (green closed circles), alternatively generated APs, but in some cases (*), two proximal dendrites independently generated Ca2+ spikes earlier than the somatic one (black closed circles). (F, G) The order of generation of Ca2+ spikes between two opposite primary dendrites and their statistics (P1 and P2 > S; both P1 and P2 Ca2+ spikes preceded the soma Ca2+ spikes; P1 > S or P2 > S, either P1 or P2 generated the earliest Ca2+ spike; S, the soma Ca2+ spikes preceded both the P1 and P2 Ca2+ spikes; n = 13). (H) Schematic models of sAP generation in multipolar DA neurons. In normal DA neurons with an attached axon, sAPs always begin in the AIS and propagate into the soma and other primary dendrites. When the AIS is disabled or removed, the excitable PDCs of many primary dendrites depolarize independently, and the one that first reaches the AP threshold fires first. Values are presented as mean ± SE. sAPs, spontaneous action potentials; PDCs, proximal dendritic compartments; AIS, axon initial segment; DA, dopamine; AP, action potential.

Fig. 4 Stochastic Ca2+ fluctuations in the PDCs and regular Ca2+ oscillations in the soma in DA neurons. (A) Fluorescence image of a dissociated DA neuron with cytosolic Ca2+ measuring sites. Ca2+ oscillations in the soma (S-black trace) and proximal dendritic region (P-red trace) were simultaneously measured with spontaneous firing. Right, mean amplitudes (red closed circle) and distributions of individual amplitudes (black open circle) of Ca2+ oscillations in the soma (0.051 ± 0.005 ∆F/F0, n = 31) and PDC (0.118 ± 0.007 ∆F/F0, n = 36). **p < 0.001. (B) Distribution of AUCs of Ca2+ oscillations in the soma (black) and PDCs (red). inset: Shaded zone indicates amount of Ca2+ influx from a single spike. (C) Coefficients of variation of Ca2+ oscillation AUCs in the proximal dendrite (SD = 0.04479, n = 36) and the soma (SD = 0.03015, n = 31). *p < 0.001. (D) Basal level of spontaneous Ca2+ oscillations in the soma and PDCs in spontaneous firing and silenced conditions by TTX treatment. Left, Ca2+ oscillation traces at the soma (black) and PDCs (red). Right, statistic results. Changes of Ca2+ levels in the soma and PDCs in the firing (proximal = 93.40 ± 1.04 µM, soma = 88.61 ± 1.96 µM, **p < 0.001, n = 6) and silenced conditions (TTX treatment, proximal = 76.98 ± 0.68 µM, soma = 78.95 ± 0.25 µM, p = 0.67, n = 6). (E) ISI variability during tonic firing was decreased by isradipine (50 µM, 0.27 ± 0.19, **p < 0.001, n = 8). (F) Dynamic interaction model between the soma and PDCs. Highly excitable and stochastically-fluctuating PDCs are electrically coupled with the regularly oscillating inertial soma. Values are presented as mean ± SE. PDCs, proximal dendritic compartments; DA, dopamine; AUCs, areas under the curves; TTX, tetrodotoxin; ISI, interspike interval; n.s., not significant.
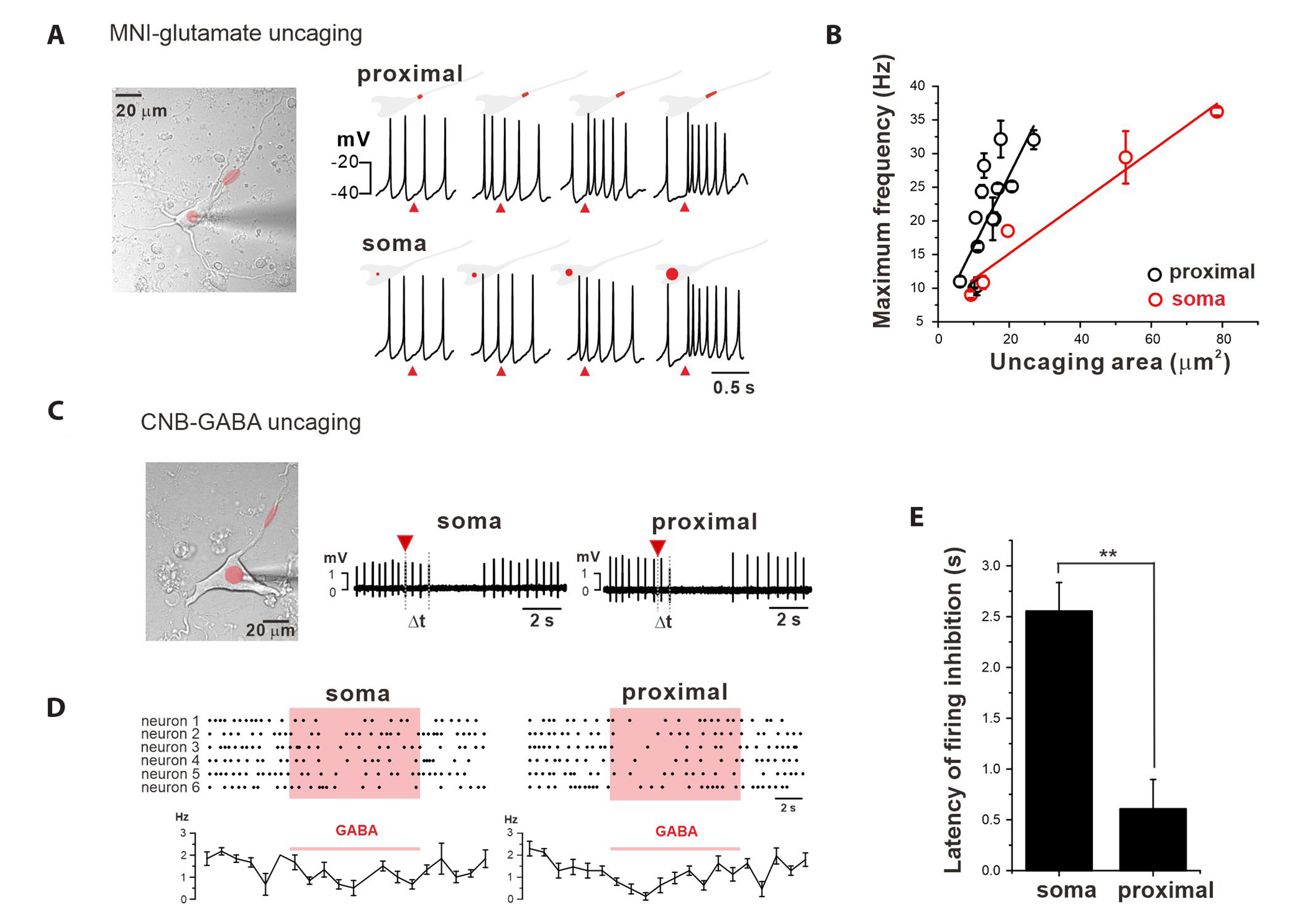
Fig. 5 PDCs are more excitable and sensitive than the soma in DA neurons. (A) A transmitted image of a dissociated DA neuron with the locations of glutamate uncaging sites (red). To get the same effect from glutamate uncaging, the soma needed a larger uncaging area than the proximal dendritic regions. (B) Relationship between the uncaged areas and the maximum firing frequencies in the soma (red) and PDCs (black). (C) A transmitted image of a dissociated DA neuron with the locations of GABA-uncaging areas (red). GABA-uncaging on the soma or proximal dendritic region induced firing inhibitions with different latency times (∆t). Red triangles indicate the uncaging times. The uncaging areas are the same. (D) GABA uncaging on the proximal dendritic region immediately inhibited spontaneous firing, in contrast with the effects on the soma. Dots indicate spikes of action potentials in DA neurons. (E) Comparison of the latency of GABA-induced firing inhibitions between the soma and PDCs (**p < 0.001, n = 6). Values are presented as mean ± SE. PDCs, proximal dendritic compartments; DA, dopamine.

Fig. 6 Dynamic coupling between the inertial soma and the excitable, stochastically-fluctuating PDCs. (A) In a spontaneously firing DA neuron, Ca2+ uncaging was performed in the soma (S) and PDCs (P). Localized Ca2+ rises were seen in the subtracted images, together with firing pauses and different Ca2+ removal kinetics between the soma and dendrite. (B) Normalized Ca2+ decays in the soma (black) and PDCs (red). (C) Half-maximal Ca2+ decay times in the soma (2.02 ± 0.15 s, n = 3) and PDCs (0.79 ± 0.003 s, n = 3). **p < 0.001. (D) Fluorescence-overlapped image of a DA neuron and a magnified dendrite image with cytosolic Ca2+ measuring sites (yellow boxes). Local Ca2+ oscillations are shown at the marked sites of a dendrite. (E) The CV of local dendritic Ca2+ oscillations increased as a function of distance from the soma. (F) Correlation coefficient of the peaks of Ca2+ oscillations (R2 = 0.67, Pearsonʼs product moment correlation coefficient, n = 5). (G) Left, A DA neuron loaded with Fluo 4-AM and Ca2+ measuring sites. Ca2+ measuring site: black circle in the soma (S), red circle in the proximal dendrite (P). Right, raw traces of Ca2+ oscillations and averaged spikes of Ca2+ oscillations in the soma and PDCs. (H) Normalized Ca2+ decay curve of soma (black) and PDCs (red) during spontaneous firing. (I) Half-maximal Ca2+ decay times in the soma (0.11 ± 0.01 s, n = 9) and PDCs (0.09 ± 0.01 s, n = 9) during spontaneous firing. Values are presented as mean ± SE. PDCs, proximal dendritic compartments; DA, dopamine; CV, coefficients of variance; n.s., not significant.
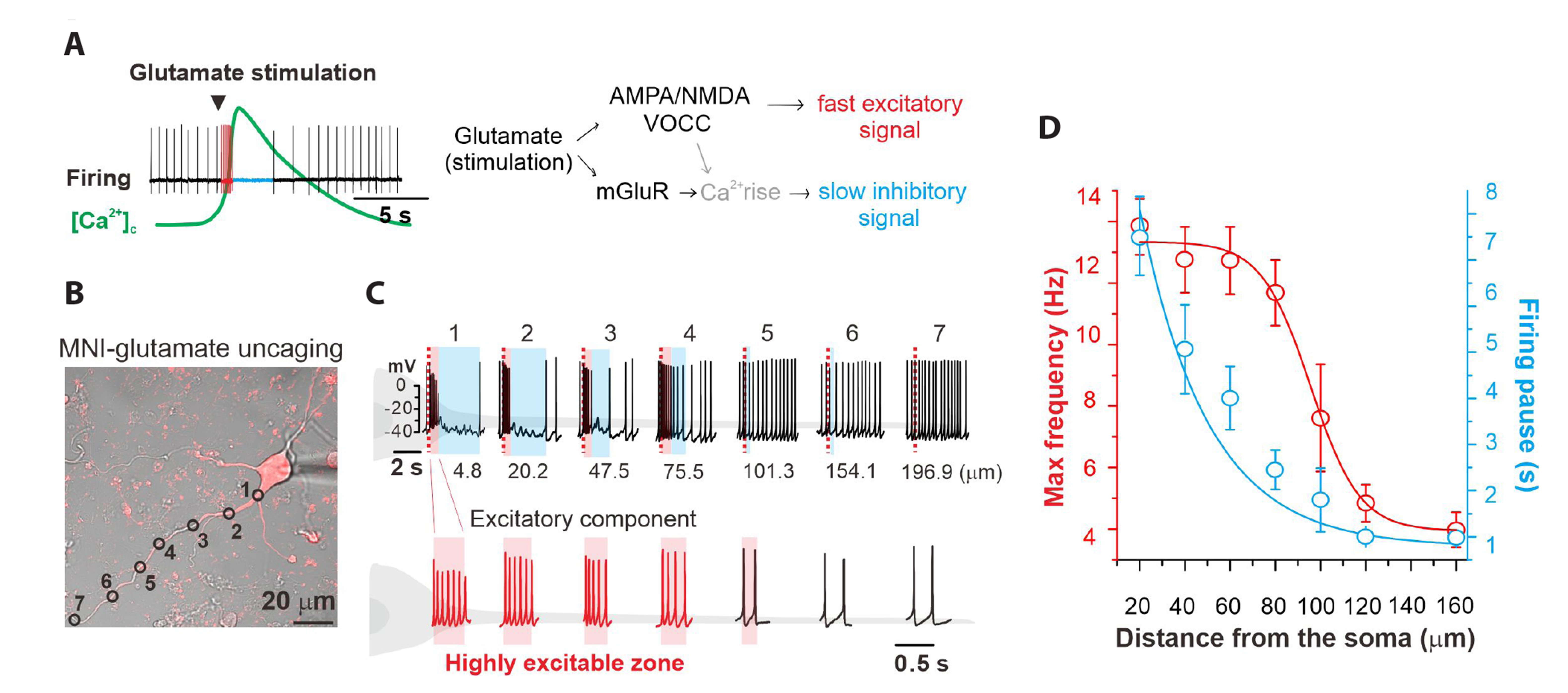
Fig. 7 Dissociation between the excitatory and inhibitory responses of spontaneous firing to glutamate in the PDCs of DA neurons. (A) Schematic drawing of glutamate signals in the DA neuron. Glutamate evoked two distinct changes in spontaneous firing and cytosolic Ca2+ levels. (B, C) In a spontaneously firing dissociated DA neuron, local glutamate uncaging was serially performed along a dendrite. Firing traces are presented according to the numbers of the marked photolysis sites and their order. Glutamate induced-firing enhancements are marked as red areas, and firing inhibitions are marked as blue areas. Bottom, expansion of firing traces in the red areas. (D) Glutamate-induced excitatory (maximum frequency, red) and inhibitory firing responses (duration of firing pause, blue) are plotted against the distance from the soma (n = 8). PDCs, proximal dendritic compartments; DA, dopamine.
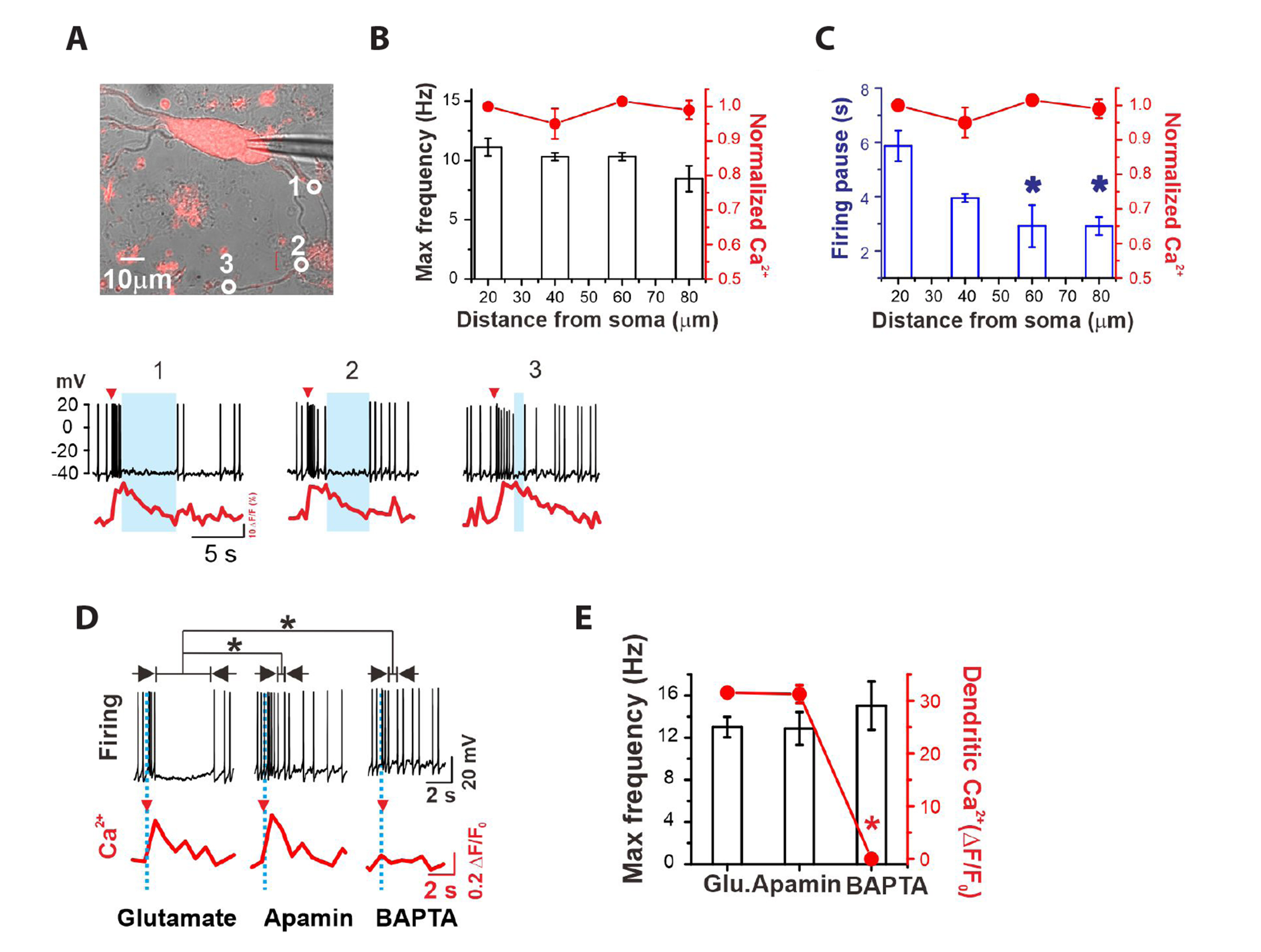
Fig. 8 Glutamate induced-firing inhibition is mediated by intracellular Ca2+ rise. (A) Simultaneous measurements of Ca2+ rises and firing inhibition upon glutamate uncaging in a dissociated dopamine neuron. Glutamate uncaging sites and Ca2+ measuring sites are marked as white circles (ø < 5 µM). Red triangles and blue boxes in the lower traces indicate uncaging points and firing pauses, respectively. (B) Local glutamate uncaging at different sites on proximal dendrites evoked Ca2+ spikes of similar amplitudes, and the evoked maximum firing frequencies did not differ significantly within the proximal dendritic regions (n = 7, *p < 0.05). (C) Postfiring pauses in spontaneous firing by glutamate uncaging in the proximal dendritic region decreased exponentially as a function of distance from the soma (n = 7). (D) Glutamate-induced firing inhibitions were blocked by apamin application (100 µM) and by dialysis with BAPTA (20 mM) using patch pipettes. The upper traces are firing changes (black), and the lower traces are dendritic Ca2+ changes (red). (E) Statistics from (D): glutamate uncaging, 13.01 ± 0.97 Hz, n = 9; apamin, 12.88 ± 1.55 Hz, n = 6; BAPTA, 15.03 ± 2.29 Hz, n = 3. Dendritic Ca2+ increases were completely inhibited by BAPTA dialysis. Values are presented as mean ± SE. *p < 0.05.

Fig. 9 The soma, responsible for inhibitory responses of spontaneous firing to glutamate, works together with the PDCs, to produce the typical burst-pause firing patterns in DA neurons. (A) A dissociated DA neuron was loaded with NP-EGTA and Fluo 4-AM. Serial photolysis of a small area along a dendrite led to localized cytosolic Ca2+ rises. Serial fluorescence images from the white box are magnified and overlapped. Dendritic Ca2+ changes in the uncaging regions are presented according to the distance from the soma, with the corresponding firing patterns recorded in the soma. Photolysis points are indicated by blue dotted lines. (B) Relative changes in the normalized hyperpolarization of membrane potential according to the distance from the soma. (C) The duration of postfiring pauses is plotted versus distance from the soma (n = 4). (D, E) When Ca2+ uncaging on the same site of a dendrite was repeated with different degrees, higher [Ca2+] rises caused longer pauses in spontaneous firing and stronger hyperpolarization of the membrane potential. Right, the relationship between the hyperpolarization of membrane potential and dendrite Ca2+ changes (n = 5, r = 0.76). (F) Strong glutamate uncaging in the local dendritic region induced different types of Ca2+ increases between the stimulated dendritic region (green) and the soma (blue). Spontaneous firing and evoked bursts were measured with a patch pipette at the soma (gray trace). (G) The recoveries of the elevated Ca2+ levels in the soma and stimulated dendritic region when the first spontaneous firing reappeared after glutamate uncaging (soma = 60.07 ± 7.35%, proximal = 2.77 ± 1.07%, n = 7). (H) Functional somatodendritic organization of the pacemaking and firing system in multipolar DA neurons. Highly excitable PDCs not only drive pacemaking but also serve as an integration site for evoked firings. PDCs, proximal dendritic compartments; DA, dopamine.
Cited by 2 articles
-
Haloperidol, a typical antipsychotic, inhibits 5-HT3 receptor-mediated currents in NCB-20 cells: a whole-cell patch-clamp study
Yong Soo Park, Gyu Min Kim, Ho Jun Sung, Ju Yeong Yu, Ki-Wug Sung
Korean J Physiol Pharmacol. 2025;29(3):349-358. doi: 10.4196/kjpp.24.320.Quetiapine competitively inhibits 5-HT3 receptor-mediated currents in NCB20 neuroblastoma cells
Yong Soo Park, Gyu Min Kim, Ho Jun Sung, Ju Yeong Yu, Ki-Wug Sung
Korean J Physiol Pharmacol. 2025;29(3):373-384. doi: 10.4196/kjpp.24.363.
Reference
-
1. Magee JC. 2000; Dendritic integration of excitatory synaptic input. Nat Rev Neurosci. 1:181–190. DOI: 10.1038/35044552. PMID: 11257906.2. Stuart GJ, Spruston N. 2015; Dendritic integration: 60 years of progress. Nat Neurosci. 18:1713–1721. DOI: 10.1038/nn.4157. PMID: 26605882.
Article3. Grace AA, Bunney BS. 1983; Intracellular and extracellular electrophysiology of nigral dopaminergic neurons--1. Identification and characterization. Neuroscience. 10:301–315. DOI: 10.1016/0306-4522(83)90135-5. PMID: 6633863.
Article4. Grace AA, Bunney BS. 1983; Intracellular and extracellular electrophysiology of nigral dopaminergic neurons--2. Action potential generating mechanisms and morphological correlates. Neuroscience. 10:317–331. DOI: 10.1016/0306-4522(83)90136-7. PMID: 6633864.
Article5. Grace AA, Onn SP. 1989; Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J Neurosci. 9:3463–3481. DOI: 10.1523/JNEUROSCI.09-10-03463.1989. PMID: 2795134. PMCID: PMC6569889.
Article6. Cossette M, Lecomte F, Parent A. 2005; Morphology and distribution of dopaminergic neurons intrinsic to the human striatum. J Chem Neuroanat. 29:1–11. DOI: 10.1016/j.jchemneu.2004.08.007. PMID: 15589697.
Article7. Häusser M, Stuart G, Racca C, Sakmann B. 1995; Axonal initiation and active dendritic propagation of action potentials in substantia nigra neurons. Neuron. 15:637–647. DOI: 10.1016/0896-6273(95)90152-3. PMID: 7546743.
Article8. Gentet LJ, Williams SR. 2007; Dopamine gates action potential backpropagation in midbrain dopaminergic neurons. J Neurosci. 27:1892–1901. DOI: 10.1523/JNEUROSCI.5234-06.2007. PMID: 17314285. PMCID: PMC6673536.
Article9. Wilson CJ, Callaway JC. 2000; Coupled oscillator model of the dopaminergic neuron of the substantia nigra. J Neurophysiol. 83:3084–3100. DOI: 10.1152/jn.2000.83.5.3084. PMID: 10805703.
Article10. Kang Y, Kitai ST. 1993; Calcium spike underlying rhythmic firing in dopaminergic neurons of the rat substantia nigra. Neurosci Res. 18:195–207. DOI: 10.1016/0168-0102(93)90055-U. PMID: 7907413.
Article11. Guzman JN, Sánchez-Padilla J, Chan CS, Surmeier DJ. 2009; Robust pacemaking in substantia nigra dopaminergic neurons. J Neurosci. 29:11011–11019. DOI: 10.1523/JNEUROSCI.2519-09.2009. PMID: 19726659. PMCID: PMC2784968.
Article12. Blythe SN, Wokosin D, Atherton JF, Bevan MD. 2009; Cellular mechanisms underlying burst firing in substantia nigra dopamine neurons. J Neurosci. 29:15531–15541. DOI: 10.1523/JNEUROSCI.2961-09.2009. PMID: 20007477. PMCID: PMC2834564.
Article13. González-Cabrera C, Meza R, Ulloa L, Merino-Sepúlveda P, Luco V, Sanhueza A, Oñate-Ponce A, Bolam JP, Henny P. 2017; Characterization of the axon initial segment of mice substantia nigra dopaminergic neurons. J Comp Neurol. 525:3529–3542. DOI: 10.1002/cne.24288. PMID: 28734032.
Article14. Jang J, Um KB, Jang M, Kim SH, Cho H, Chung S, Kim HJ, Park MK. 2014; Balance between the proximal dendritic compartment and the soma determines spontaneous firing rate in midbrain dopamine neurons. J Physiol. 592:2829–2844. DOI: 10.1113/jphysiol.2014.275032. PMID: 24756642. PMCID: PMC4221823.
Article15. Medvedev GS, Wilson CJ, Callaway JC, Kopell N. 2003; Dendritic synchrony and transient dynamics in a coupled oscillator model of the dopaminergic neuron. J Comput Neurosci. 15:53–69. DOI: 10.1023/A:1024422802673. PMID: 12843695.16. Grace AA, Floresco SB, Goto Y, Lodge DJ. 2007; Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci. 30:220–227. DOI: 10.1016/j.tins.2007.03.003. PMID: 17400299.
Article17. Pucak ML, Grace AA. 1994; Regulation of substantia nigra dopamine neurons. Crit Rev Neurobiol. 9:67–89.18. Overton PG, Clark D. 1997; Burst firing in midbrain dopaminergic neurons. Brain Res Brain Res Rev. 25:312–334. DOI: 10.1016/S0165-0173(97)00039-8. PMID: 9495561.
Article19. Hyland BI, Reynolds JN, Hay J, Perk CG, Miller R. 2002; Firing modes of midbrain dopamine cells in the freely moving rat. Neuroscience. 114:475–492. DOI: 10.1016/S0306-4522(02)00267-1. PMID: 12204216.
Article20. Tsai HC, Zhang F, Adamantidis A, Stuber GD, Bonci A, de Lecea L, Deisseroth K. 2009; Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning. Science. 324:1080–1084. DOI: 10.1126/science.1168878. PMID: 19389999. PMCID: PMC5262197.
Article21. Wise RA. 2009; Roles for nigrostriatal--not just mesocorticolimbic--dopamine in reward and addiction. Trends Neurosci. 32:517–524. DOI: 10.1016/j.tins.2009.06.004. PMID: 19758714. PMCID: PMC2755633.
Article22. Dawson TM, Dawson VL. 2003; Molecular pathways of neurodegeneration in Parkinsonʼs disease. Science. 302:819–822. DOI: 10.1126/science.1087753. PMID: 14593166.
Article23. Surmeier DJ, Guzman JN, Sanchez J, Schumacker PT. 2012; Physiological phenotype and vulnerability in Parkinsonʼs disease. Cold Spring Harb Perspect Med. 2:a009290. DOI: 10.1101/cshperspect.a009290. PMID: 22762023. PMCID: PMC3385938.
Article24. Grace AA. 1988; In vivo and in vitro intracellular recordings from rat midbrain dopamine neurons. Ann N Y Acad Sci. 537:51–76. DOI: 10.1111/j.1749-6632.1988.tb42096.x. PMID: 2849358.
Article25. Um KB, Hahn S, Kim SW, Lee YJ, Birnbaumer L, Kim HJ, Park MK. 2021; TRPC3 and NALCN channels drive pacemaking in substantia nigra dopaminergic neurons. Elife. 10:e70920. DOI: 10.7554/eLife.70920. PMID: 34409942. PMCID: PMC8456572.
Article26. Hahn S, Um KB, Kim SW, Kim HJ, Park MK. 2023; Proximal dendritic localization of NALCN channels underlies tonic and burst firing in nigral dopaminergic neurons. J Physiol. 601:171–193. DOI: 10.1113/JP283716. PMID: 36398712.
Article27. Jang M, Um KB, Jang J, Kim HJ, Cho H, Chung S, Park MK. 2015; Coexistence of glutamatergic spine synapses and shaft synapses in substantia nigra dopamine neurons. Sci Rep. 5:14773. DOI: 10.1038/srep14773. PMID: 26435058. PMCID: PMC4593176.
Article28. Choi YM, Kim SH, Uhm DY, Park MK. 2003; Glutamate-mediated [Ca2+]c dynamics in spontaneously firing dopamine neurons of the rat substantia nigra pars compacta. J Cell Sci. 116:2665–2675. DOI: 10.1242/jcs.00481. PMID: 12746490.
Article29. Hahn J, Tse TE, Levitan ES. 2003; Long-term K+ channel-mediated dampening of dopamine neuron excitability by the antipsychotic drug haloperidol. J Neurosci. 23:10859–10866. DOI: 10.1523/JNEUROSCI.23-34-10859.2003. PMID: 14645479. PMCID: PMC6740975.
Article30. Ping HX, Shepard PD. 1999; Blockade of SK-type Ca2+-activated K+ channels uncovers a Ca2+-dependent slow afterdepolarization in nigral dopamine neurons. J Neurophysiol. 81:977–984. DOI: 10.1152/jn.1999.81.3.977. PMID: 10085326.
Article31. Wolfart J, Roeper J. 2002; Selective coupling of T-type calcium channels to SK potassium channels prevents intrinsic bursting in dopaminergic midbrain neurons. J Neurosci. 22:3404–3413. Erratum in: J Neurosci. 2002;22:5250. DOI: 10.1523/JNEUROSCI.22-09-03404.2002. PMID: 11978817. PMCID: PMC6758365.
Article32. Wolfart J, Neuhoff H, Franz O, Roeper J. 2001; Differential expression of the small-conductance, calcium-activated potassium channel SK3 is critical for pacemaker control in dopaminergic midbrain neurons. J Neurosci. 21:3443–3456. DOI: 10.1523/JNEUROSCI.21-10-03443.2001. PMID: 11331374. PMCID: PMC6762487.
Article33. Deignan J, Luján R, Bond C, Riegel A, Watanabe M, Williams JT, Maylie J, Adelman JP. 2012; SK2 and SK3 expression differentially affect firing frequency and precision in dopamine neurons. Neuroscience. 217:67–76. DOI: 10.1016/j.neuroscience.2012.04.053. PMID: 22554781. PMCID: PMC3383402.
Article34. Kuznetsov AS, Kopell NJ, Wilson CJ. 2006; Transient high-frequency firing in a coupled-oscillator model of the mesencephalic dopaminergic neuron. J Neurophysiol. 95:932–947. DOI: 10.1152/jn.00691.2004. PMID: 16207783.
Article35. Chuhma N, Zhang H, Masson J, Zhuang X, Sulzer D, Hen R, Rayport S. 2004; Dopamine neurons mediate a fast excitatory signal via their glutamatergic synapses. J Neurosci. 24:972–981. DOI: 10.1523/JNEUROSCI.4317-03.2004. PMID: 14749442. PMCID: PMC6729804.
Article36. Fiorillo CD, Williams JT. 1998; Glutamate mediates an inhibitory postsynaptic potential in dopamine neurons. Nature. 394:78–82. DOI: 10.1038/27919. PMID: 9665131.
Article37. Kim SH, Choi YM, Chung S, Uhm DY, Park MK. 2004; Two different Ca2+-dependent inhibitory mechanisms of spontaneous firing by glutamate in dopamine neurons. J Neurochem. 91:983–995. DOI: 10.1111/j.1471-4159.2004.02783.x. PMID: 15525352.
Article38. Morikawa H, Imani F, Khodakhah K, Williams JT. 2000; Inositol 1,4,5-triphosphate-evoked responses in midbrain dopamine neurons. J Neurosci. 20:RC103. DOI: 10.1523/JNEUROSCI.20-20-j0003.2000. PMID: 11027254. PMCID: PMC6772861.
Article39. Berridge MJ, Bootman MD, Roderick HL. 2003; Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 4:517–529. DOI: 10.1038/nrm1155. PMID: 12838335.
Article40. Papaioannou VE, Verkerk AO, Amin AS, de Bakker JM. 2013; Intracardiac origin of heart rate variability, pacemaker funny current and their possible association with critical illness. Curr Cardiol Rev. 9:82–96. DOI: 10.2174/1573403X11309010010.
Article41. Yaniv Y, Lyashkov AE, Sirenko S, Okamoto Y, Guiriba TR, Ziman BD, Morrell CH, Lakatta EG. 2014; Stochasticity intrinsic to coupled-clock mechanisms underlies beat-to-beat variability of spontaneous action potential firing in sinoatrial node pacemaker cells. J Mol Cell Cardiol. 77:1–10. DOI: 10.1016/j.yjmcc.2014.09.008. PMID: 25257916. PMCID: PMC4312254.
Article42. Anwar H, Hepburn I, Nedelescu H, Chen W, De Schutter E. 2013; Stochastic calcium mechanisms cause dendritic calcium spike variability. J Neurosci. 33:15848–15867. DOI: 10.1523/JNEUROSCI.1722-13.2013. PMID: 24089492. PMCID: PMC6618479.
Article43. Deister CA, Teagarden MA, Wilson CJ, Paladini CA. 2009; An intrinsic neuronal oscillator underlies dopaminergic neuron bursting. J Neurosci. 29:15888–15897. DOI: 10.1523/JNEUROSCI.4053-09.2009. PMID: 20016105. PMCID: PMC2824818.
Article44. Kuznetsova AY, Huertas MA, Kuznetsov AS, Paladini CA, Canavier CC. 2010; Regulation of firing frequency in a computational model of a midbrain dopaminergic neuron. J Comput Neurosci. 28:389–403. DOI: 10.1007/s10827-010-0222-y. PMID: 20217204. PMCID: PMC2929809.
Article45. Kole MH, Ilschner SU, Kampa BM, Williams SR, Ruben PC, Stuart GJ. 2008; Action potential generation requires a high sodium channel density in the axon initial segment. Nat Neurosci. 11:178–186. DOI: 10.1038/nn2040. PMID: 18204443.
Article46. Debanne D, Campanac E, Bialowas A, Carlier E, Alcaraz G. 2011; Axon physiology. Physiol Rev. 91:555–602. DOI: 10.1152/physrev.00048.2009. PMID: 21527732.
Article47. Tucker KR, Huertas MA, Horn JP, Canavier CC, Levitan ES. 2012; Pacemaker rate and depolarization block in nigral dopamine neurons: a somatic sodium channel balancing act. J Neurosci. 32:14519–14531. DOI: 10.1523/JNEUROSCI.1251-12.2012. PMID: 23077037. PMCID: PMC3494994.
Article48. Rall W, Burke RE, Holmes WR, Jack JJ, Redman SJ, Segev I. 1992; Matching dendritic neuron models to experimental data. Physiol Rev. 72(4 Suppl):S159–S186. DOI: 10.1152/physrev.1992.72.suppl_4.S159. PMID: 1438585.
Article49. Migliore M, Shepherd GM. 2002; Emerging rules for the distributions of active dendritic conductances. Nat Rev Neurosci. 3:362–370. DOI: 10.1038/nrn810. PMID: 11988775.
Article50. Amini B, Clark JW Jr, Canavier CC. 1999; Calcium dynamics underlying pacemaker-like and burst firing oscillations in midbrain dopaminergic neurons: a computational study. J Neurophysiol. 82:2249–2261. DOI: 10.1152/jn.1999.82.5.2249. PMID: 10561403.
Article51. Katayama J, Akaike N, Nabekura J. 2003; Characterization of pre- and post-synaptic metabotropic glutamate receptor-mediated inhibitory responses in substantia nigra dopamine neurons. Neurosci Res. 45:101–115. DOI: 10.1016/S0168-0102(02)00202-X. PMID: 12507729.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Regional difference in spontaneous firing inhibition by GABA(A) and GABA(B) receptors in nigral dopamine neurons
- In vitro generation of mature midbrain-type dopamine neurons by adjusting exogenous Nurr1 and Foxa2 expressions to their physiologic patterns
- Study on the Effect of Carbon Monoxide Intoxication to Serotonergic Neurons in Midbrain Raphe Nuclei of the Rat
- The Effect of Treatment with Tryptophan and/or Imipramine on the Serotonergic Immunoreactivity in Raphe Nucleus of Midbrain of the Rats
- Effects of 6-hydroxydopamine on the Adult Neurogenesis of Dopaminergic Neurons in the Mouse Midbrain
