The Link between Mitochondrial Dysfunction and Sarcopenia: An Update Focusing on the Role of Pyruvate Dehydrogenase Kinase 4
- Affiliations
-
- 1Department of Internal Medicine, Kyungpook National University Chilgok Hospital, School of Medicine, Kyungpook National University, Daegu, Korea
- 2Bio-Medical Research Institute, Kyungpook National University Hospital, Daegu, Korea
- 3Department of Biomedical Science, Graduate School, Kyungpook National University, Daegu, Korea
- 4BK21 Plus KNU Biomedical Convergence Program, Kyungpook National University, Daegu, Korea
- 5Department of Internal Medicine, Kyungpook National University Hospital, School of Medicine, Kyungpook National University, Daegu, Korea
- KMID: 2540512
- DOI: http://doi.org/10.4093/dmj.2022.0305
Abstract
- Sarcopenia, defined as a progressive loss of muscle mass and function, is typified by mitochondrial dysfunction and loss of mitochondrial resilience. Sarcopenia is associated not only with aging, but also with various metabolic diseases characterized by mitochondrial dyshomeostasis. Pyruvate dehydrogenase kinases (PDKs) are mitochondrial enzymes that inhibit the pyruvate dehydrogenase complex, which controls pyruvate entry into the tricarboxylic acid cycle and the subsequent adenosine triphosphate production required for normal cellular activities. PDK4 is upregulated in mitochondrial dysfunction-related metabolic diseases, especially pathologic muscle conditions associated with enhanced muscle proteolysis and aberrant myogenesis. Increases in PDK4 are associated with perturbation of mitochondria-associated membranes and mitochondrial quality control, which are emerging as a central mechanism in the pathogenesis of metabolic disease-associated muscle atrophy. Here, we review how mitochondrial dysfunction affects sarcopenia, focusing on the role of PDK4 in mitochondrial homeostasis. We discuss the molecular mechanisms underlying the effects of PDK4 on mitochondrial dysfunction in sarcopenia and show that targeting mitochondria could be a therapeutic target for treating sarcopenia.
Keyword
Figure
-
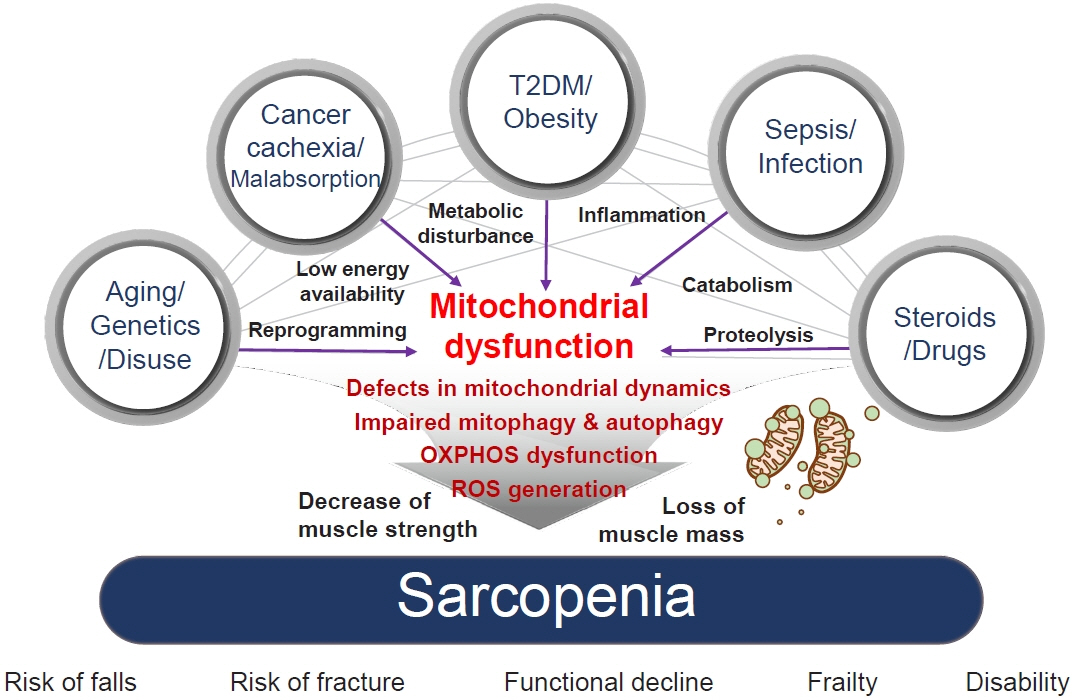
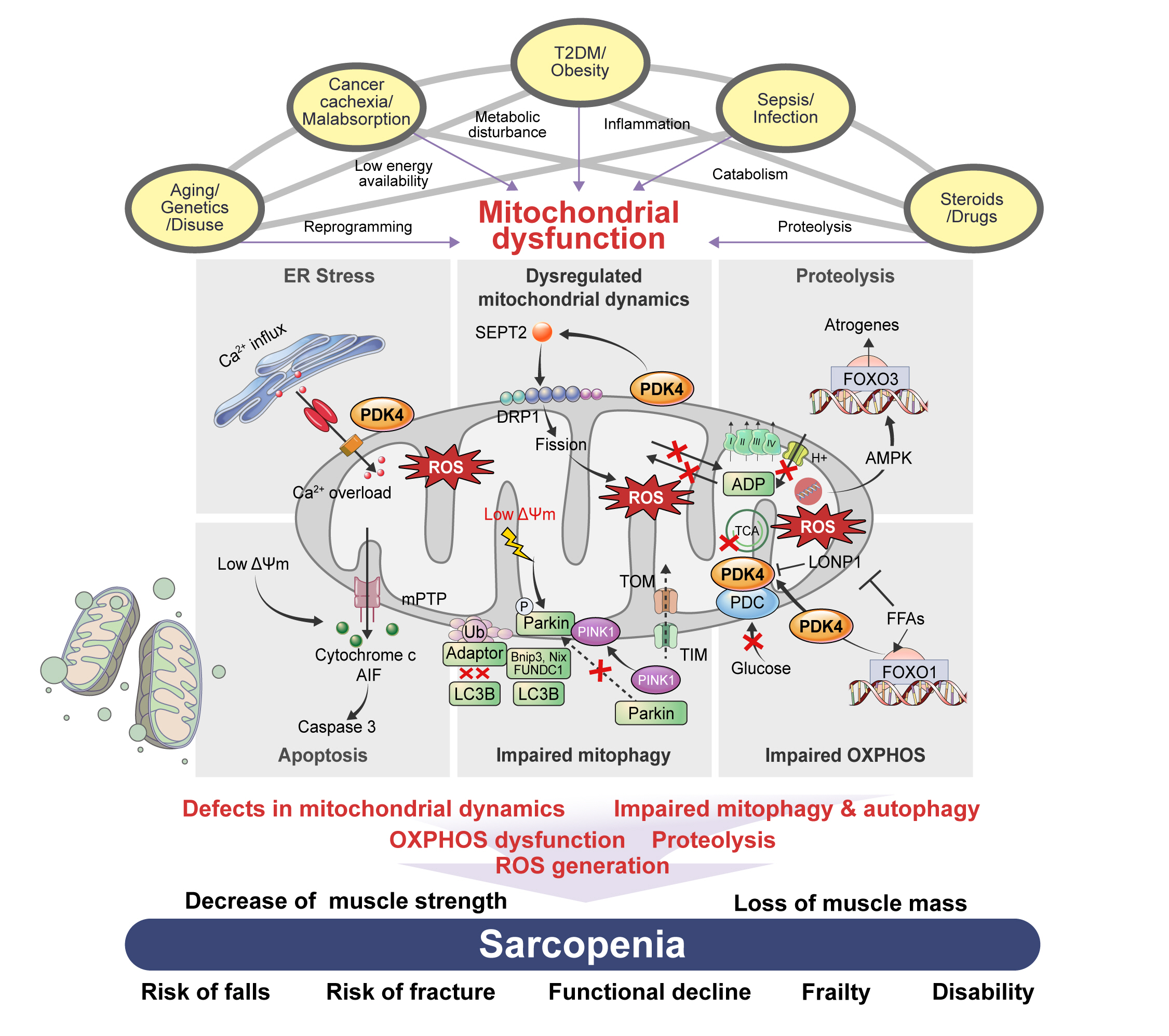
Fig. 1. The association between risk factors of sarcopenia and mitochondrial dysfunction. Sarcopenia, generally defined as aging-related loss of muscle mass and function, is closely associated with other genetic or environmental factors. Sarcopenia pathogenesis occurs in multiple ways: genetic predisposition, aging, and disuse of muscle lead to transcriptional reprogramming or epigenetic modification; cancer cachexia or nutrient malabsorption cause low energy availability; metabolic diseases, such as type 2 diabetes mellitus (T2DM) and obesity, lead to metabolic disturbances; infection and sepsis cause inflammation; and steroids and other drugs can lead to catabolism and proteolysis. All these risk factors cause mitochondrial dysfunction, which results in sarcopenia via several mechanisms: defects in mitochondrial dynamics, such as fusion/fission; impaired mitochondrial quality control, including mitochondrial biogenesis; mitophagy and autophagy; dysfunctions in oxidative phosphorylation (OXPHOS), which is important in energy generation; and reactive oxygen species (ROS) production, which originates in dysregulated OXPHOS. Mechanistically, these factors decrease muscle strength and cause loss of muscle mass, which are associated with an increased risk of falls and fractures, functional decline, frailty, disability, and hospitalization with poorer health outcomes.
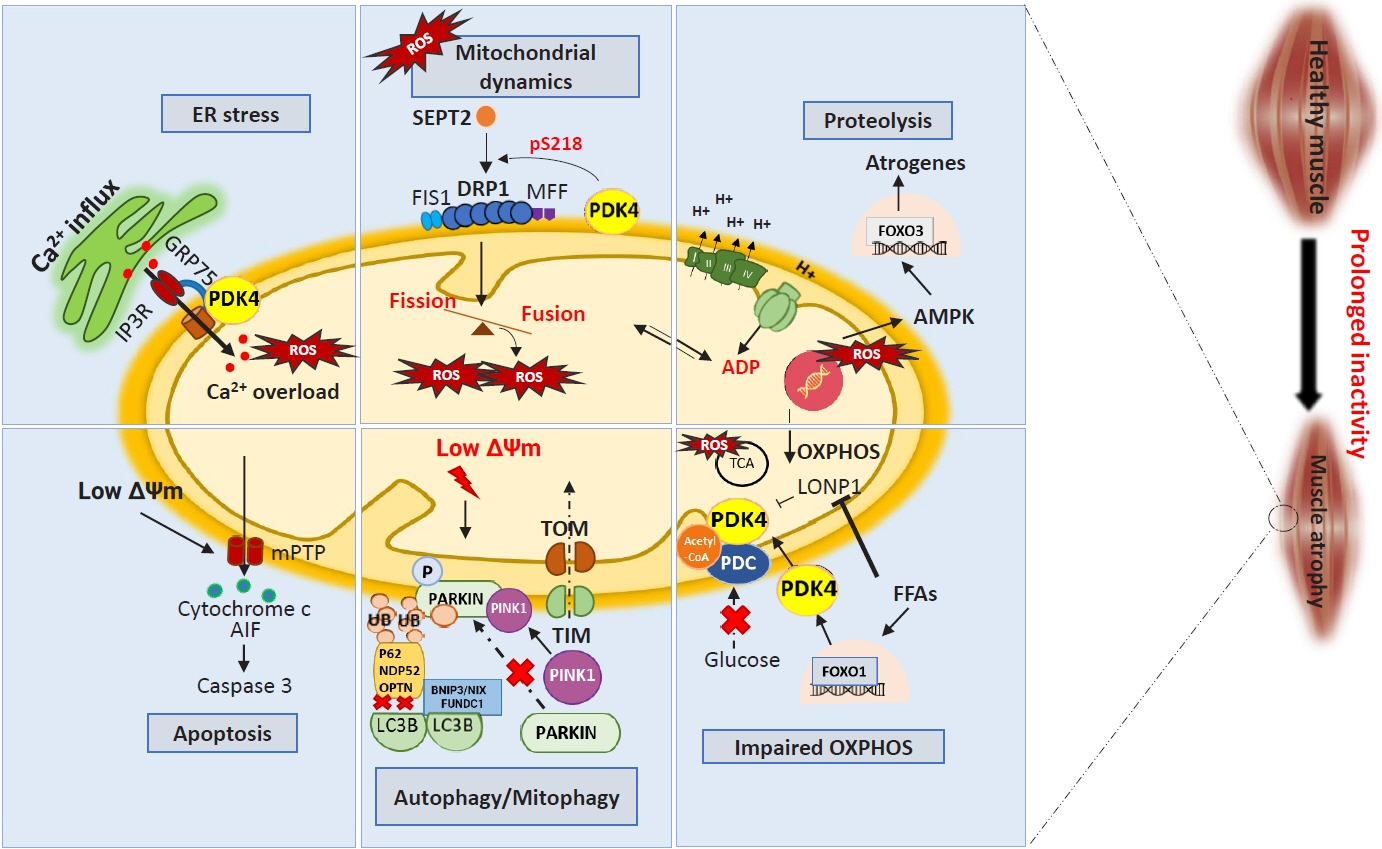
Fig. 2. Muscle atrophy induces mitochondrial dysfunction through multiple pathways. Prolonged inactivity and other myopathies elevate reactive oxygen species (ROS) levels, which regulate different mitochondrial systems. Increased endoplasmic reticulum (ER) stress caused by calcium (Ca2+) overload and ROS results in Ca2+ influx into mitochondria through pyruvate dehydrogenase kinase 4 (PDK4)-mediated stabilization of inositol 1,4,5 trisphosphate receptor type 1 (IP3R1)–glucose-regulated protein 75 (GRP75)–voltage-dependent anion-selective channel 1 complex at the mitochondria-associated ER membrane. Increased mitochondrial Ca2+ content causes a drop in mitochondrial membrane potential (ΔΨm) leading to the opening of mitochondrial permeability transition pores (mPTP) to release cytochrome c, which activates apoptosis-inducing factor (AIF), leading to the onset of caspase 3-mediated cell death. Muscle atrophy causes increased fission via septin 2 (SEPT2)-mediated dynamin-related protein 1 (DRP1) upregulation. This imbalance between fusion and fission generates ROS and a lower ΔΨm, leading to dysfunctional mitochondrial-mediated autophagy (mitophagy). This occurs via suppressed PTEN-induced kinase 1 (PINK1)-PARKIN interaction, as well as reduced autophagic flux by lysosome adapter accumulation. Fission also causes a drop in ATP synthesis and impaired electron transport chain activity, leading to the activation of 5´ AMP-activated protein kinase (AMPK)–forkhead box protein O3 (FOXO3)-dependent atrogenes, which cause protein degradation (proteolysis). An increase in free-fatty acids (FFAs) caused by myopathologic metabolic alterations results in a switch from glucose oxidation to beta-oxidation. This causes an increase in FOXO1-PDK4 activity leading to the inactivation of the pyruvate dehydrogenase complex (PDC) by PDK4. Synergistically, FFAs also inhibit Lon peptidase 1 (LonP1), which degrades PDK4 in the mitochondria. Overall, this decreases oxidative phosphorylation (OXPHOS), leading to metabolic inflexibility-related myopathies. Acetyl-CoA, acetyl coenzyme A; ADP, adenosine diphosphate; BNIP3, BCL2/adenovirus E1B 19 kDa protein-interacting protein 3; FIS1, mitochondrial fission 1 protein; FUNDC1, FUN14 domain containing 1; H+, proton ion; LC3B, microtubule-associated proteins 1A/1B light chain 3B; MFF, mitochondrial fission factor; NDP52, nuclear dot protein 52; OPTN, optineurin; P62, sequestosome 1 (SQSTM1); TCA, tricarboxylic acid cycle; TIM, translocase of the inner membrane; TOM, translocase of the outer membrane; Ub, ubiquitin.
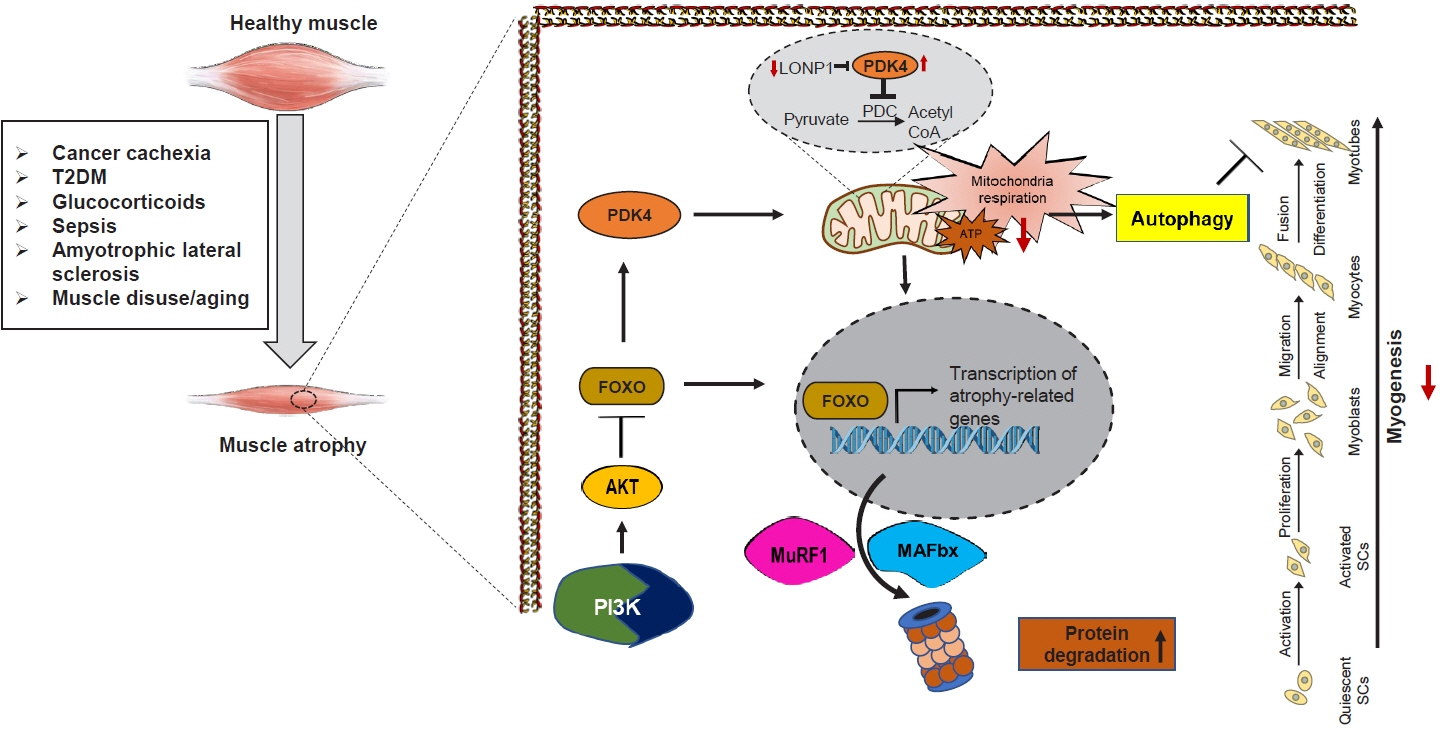
Fig. 3. Schematic of the involvement of pyruvate dehydrogenase kinase 4 (PDK4) in regulating muscle protein degradation and atrophic gene expression. In the pathological state, PDK4 is activated and can contribute to mitochondrial damage, as well as transcription of atrophy-related genes culminating in muscle atrophy. The transcription factor forkhead box protein O1 (FOXO1) is proposed to upregulate transcription of PDK4. During starvation, a decline in insulin levels activates FOXO1, which enhances PDK4 expression. PDK4 is highly expressed in cancer cachexia, type 2 diabetes mellitus (T2DM), glucocorticoid-induced myopathy, sepsis, amyotrophic lateral sclerosis (ALS), muscle disuse, and aging. Overexpression of PDK4 induces mitochondrial dysfunction and muscle protein degradation, giving PDK4 a central role in mediating metabolic instability-induced skeletal muscle dysfunction. Acetyl-CoA, acetyl coenzyme A; ATP, adenosine triphosphate; LonP1, Lon peptidase 1; MAFbx, muscle atrophy F-box; MuRF1, muscle RING finger 1; PDC, pyruvate dehydrogenase complex; PI3K, phosphatidylinositol-3-kinase; SC, satellite cell.

Fig. 4. Pyruvate dehydrogenase kinase 4 (PDK4) and regulation of myogenesis. The ubiquitin-proteasome pathway serves a pivotal role in the mediation of protein degradation in muscle atrophy. In normal muscle, the transcription factor myogenin (MYOG) activates transcription of the myogenic genes MYMX (myomixer, myoblast fusion factor) and MYMK (myomaker, myoblast fusion factor). By contrast, during dexamethasone-induced muscle atrophy, induction of PDK4 leads to phosphorylation (P) of myogenin (MYOG) and recruitment of muscle atrophy F-box (MAFbx), which polyubiquitinates (Ub) MYOG, leading to its degradation and preventing transcription of myogenic genes. MyHC, myosin heavy chain; UPS, ubiquitin-proteasome system.
Reference
-
1. Shimokata H, Shimada H, Satake S, Endo N, Shibasaki K, Ogawa S, et al. Chapter 2 Epidemiology of sarcopenia. Geriatr Gerontol Int. 2018; 18 Suppl 1:13–22.2. Fu X, Zhu M, Zhang S, Foretz M, Viollet B, Du M. Obesity impairs skeletal muscle regeneration through inhibition of AMPK. Diabetes. 2016; 65:188–200.3. Liu X, Qu H, Zheng Y, Liao Q, Zhang L, Liao X, et al. Mitochondrial glycerol 3-phosphate dehydrogenase promotes skeletal muscle regeneration. EMBO Mol Med. 2018; 10:e9390.4. Hyatt H, Deminice R, Yoshihara T, Powers SK. Mitochondrial dysfunction induces muscle atrophy during prolonged inactivity: a review of the causes and effects. Arch Biochem Biophys. 2019; 662:49–60.5. Pin F, Novinger LJ, Huot JR, Harris RA, Couch ME, O’Connell TM, et al. PDK4 drives metabolic alterations and muscle atrophy in cancer cachexia. FASEB J. 2019; 33:7778–90.6. Abbatecola AM, Paolisso G, Fattoretti P, Evans WJ, Fiore V, Dicioccio L, et al. Discovering pathways of sarcopenia in older adults: a role for insulin resistance on mitochondria dysfunction. J Nutr Health Aging. 2011; 15:890–5.7. Picca A, Calvani R, Bossola M, Allocca E, Menghi A, Pesce V, et al. Update on mitochondria and muscle aging: all wrong roads lead to sarcopenia. Biol Chem. 2018; 399:421–36.8. Barbieri E, Sestili P. Reactive oxygen species in skeletal muscle signaling. J Signal Transduct. 2012; 2012:982794.9. Boncompagni S, Rossi AE, Micaroni M, Beznoussenko GV, Polishchuk RS, Dirksen RT, et al. Mitochondria are linked to calcium stores in striated muscle by developmentally regulated tethering structures. Mol Biol Cell. 2009; 20:1058–67.10. Twig G, Hyde B, Shirihai OS. Mitochondrial fusion, fission and autophagy as a quality control axis: the bioenergetic view. Biochim Biophys Acta. 2008; 1777:1092–7.11. Zhang S, Hulver MW, McMillan RP, Cline MA, Gilbert ER. The pivotal role of pyruvate dehydrogenase kinases in metabolic flexibility. Nutr Metab (Lond). 2014; 11:10.12. Bowker-Kinley MM, Davis WI, Wu P, Harris RA, Popov KM. Evidence for existence of tissue-specific regulation of the mammalian pyruvate dehydrogenase complex. Biochem J. 1998; 329(Pt 1):191–6.13. Roche TE, Hiromasa Y. Pyruvate dehydrogenase kinase regulatory mechanisms and inhibition in treating diabetes, heart ischemia, and cancer. Cell Mol Life Sci. 2007; 64:830–49.14. Sugden MC, Holness MJ. Mechanisms underlying regulation of the expression and activities of the mammalian pyruvate dehydrogenase kinases. Arch Physiol Biochem. 2006; 112:139–49.15. Park S, Jeon JH, Min BK, Ha CM, Thoudam T, Park BY, et al. Role of the pyruvate dehydrogenase complex in metabolic remodeling: differential pyruvate dehydrogenase complex functions in metabolism. Diabetes Metab J. 2018; 42:270–81.16. Holness MJ, Kraus A, Harris RA, Sugden MC. Targeted upregulation of pyruvate dehydrogenase kinase (PDK)-4 in slowtwitch skeletal muscle underlies the stable modification of the regulatory characteristics of PDK induced by high-fat feeding. Diabetes. 2000; 49:775–81.17. Crossland H, Constantin-Teodosiu D, Greenhaff PL, Gardiner SM. Low-dose dexamethasone prevents endotoxaemia-induced muscle protein loss and impairment of carbohydrate oxidation in rat skeletal muscle. J Physiol. 2010; 588(Pt 8):1333–47.18. Lee IK. The role of pyruvate dehydrogenase kinase in diabetes and obesity. Diabetes Metab J. 2014; 38:181–6.19. Manickam R, Oh HY, Tan CK, Paramalingam E, Wahli W. Metronidazole causes skeletal muscle atrophy and modulates muscle chronometabolism. Int J Mol Sci. 2018; 19:2418.20. Zhang L, Keung W, Samokhvalov V, Wang W, Lopaschuk GD. Role of fatty acid uptake and fatty acid beta-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim Biophys Acta. 2010; 1801:1–22.21. Jeon JH, Thoudam T, Choi EJ, Kim MJ, Harris RA, Lee IK. Loss of metabolic flexibility as a result of overexpression of pyruvate dehydrogenase kinases in muscle, liver and the immune system: therapeutic targets in metabolic diseases. J Diabetes Investig. 2021; 12:21–31.22. Thoudam T, Ha CM, Leem J, Chanda D, Park JS, Kim HJ, et al. PDK4 augments ER-mitochondria contact to dampen skeletal muscle insulin signaling during obesity. Diabetes. 2019; 68:571–86.23. Calvani R, Joseph AM, Adhihetty PJ, Miccheli A, Bossola M, Leeuwenburgh C, et al. Mitochondrial pathways in sarcopenia of aging and disuse muscle atrophy. Biol Chem. 2013; 394:393–414.24. Anderson EJ, Lustig ME, Boyle KE, Woodlief TL, Kane DA, Lin CT, et al. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J Clin Invest. 2009; 119:573–81.25. Palamiuc L, Schlagowski A, Ngo ST, Vernay A, Dirrig-Grosch S, Henriques A, et al. A metabolic switch toward lipid use in glycolytic muscle is an early pathologic event in a mouse model of amyotrophic lateral sclerosis. EMBO Mol Med. 2015; 7:526–46.26. Nahle Z, Hsieh M, Pietka T, Coburn CT, Grimaldi PA, Zhang MQ, et al. CD36-dependent regulation of muscle FoxO1 and PDK4 in the PPAR delta/beta-mediated adaptation to metabolic stress. J Biol Chem. 2008; 283:14317–26.27. Yan D, Cai Y, Luo J, Liu J, Li X, Ying F, et al. FOXO1 contributes to diabetic cardiomyopathy via inducing imbalanced oxidative metabolism in type 1 diabetes. J Cell Mol Med. 2020; 24:7850–61.28. Xu Z, Fu T, Guo Q, Zhou D, Sun W, Zhou Z, et al. Disuse-associated loss of the protease LONP1 in muscle impairs mitochondrial function and causes reduced skeletal muscle mass and strength. Nat Commun. 2022; 13:894.29. Sanchez AM, Csibi A, Raibon A, Cornille K, Gay S, Bernardi H, et al. AMPK promotes skeletal muscle autophagy through activation of forkhead FoxO3a and interaction with Ulk1. J Cell Biochem. 2012; 113:695–710.30. Crewe C, Schafer C, Lee I, Kinter M, Szweda LI. Regulation of pyruvate dehydrogenase kinase 4 in the heart through degradation by the lon protease in response to mitochondrial substrate availability. J Biol Chem. 2017; 292:305–12.31. Rossi A, Pizzo P, Filadi R. Calcium, mitochondria and cell metabolism: a functional triangle in bioenergetics. Biochim Biophys Acta Mol Cell Res. 2019; 1866:1068–78.32. Pauly M, Angebault-Prouteau C, Dridi H, Notarnicola C, Scheuermann V, Lacampagne A, et al. ER stress disturbs SR/ ER-mitochondria Ca2+ transfer: implications in Duchenne muscular dystrophy. Biochim Biophys Acta Mol Basis Dis. 2017; 1863:2229–39.33. Arruda AP, Hotamisligil GS. Calcium homeostasis and organelle function in the pathogenesis of obesity and diabetes. Cell Metab. 2015; 22:381–97.34. Romanello V, Guadagnin E, Gomes L, Roder I, Sandri C, Petersen Y, et al. Mitochondrial fission and remodelling contributes to muscle atrophy. EMBO J. 2010; 29:1774–85.35. Xu C, Kasimumali A, Guo X, Lu R, Xie K, Zhu M, et al. Reduction of mitochondria and up regulation of pyruvate dehydrogenase kinase 4 of skeletal muscle in patients with chronic kidney disease. Nephrology (Carlton). 2020; 25:230–8.36. Thoudam T, Chanda D, Sinam IS, Kim BG, Kim MJ, Oh CJ, et al. Noncanonical PDK4 action alters mitochondrial dynamics to affect the cellular respiratory status. Proc Natl Acad Sci U S A. 2022; 119:e2120157119.37. Leduc-Gaudet JP, Hussain SN, Barreiro E, Gouspillou G. Mitochondrial dynamics and mitophagy in skeletal muscle health and aging. Int J Mol Sci. 2021; 22:8179.38. Drummond MJ, Addison O, Brunker L, Hopkins PN, McClain DA, LaStayo PC, et al. Downregulation of E3 ubiquitin ligases and mitophagy-related genes in skeletal muscle of physically inactive, frail older women: a cross-sectional comparison. J Gerontol A Biol Sci Med Sci. 2014; 69:1040–8.39. Chen CC, Erlich AT, Hood DA. Role of Parkin and endurance training on mitochondrial turnover in skeletal muscle. Skelet Muscle. 2018; 8:10.40. Sebastian D, Sorianello E, Segales J, Irazoki A, Ruiz-Bonilla V, Sala D, et al. Mfn2 deficiency links age-related sarcopenia and impaired autophagy to activation of an adaptive mitophagy pathway. EMBO J. 2016; 35:1677–93.41. Park S, Choi SG, Yoo SM, Nah J, Jeong E, Kim H, et al. Pyruvate stimulates mitophagy via PINK1 stabilization. Cell Signal. 2015; 27:1824–30.42. Peters SJ, Harris RA, Heigenhauser GJ, Spriet LL. Muscle fiber type comparison of PDH kinase activity and isoform expression in fed and fasted rats. Am J Physiol Regul Integr Comp Physiol. 2001; 280:R661–8.43. Furuyama T, Kitayama K, Yamashita H, Mori N. Forkhead transcription factor FOXO1 (FKHR)-dependent induction of PDK4 gene expression in skeletal muscle during energy deprivation. Biochem J. 2003; 375(Pt 2):365–71.44. Wu P, Peters JM, Harris RA. Adaptive increase in pyruvate dehydrogenase kinase 4 during starvation is mediated by peroxisome proliferator-activated receptor alpha. Biochem Biophys Res Commun. 2001; 287:391–6.45. Wende AR, Huss JM, Schaeffer PJ, Giguere V, Kelly DP. PGC1alpha coactivates PDK4 gene expression via the orphan nuclear receptor ERRalpha: a mechanism for transcriptional control of muscle glucose metabolism. Mol Cell Biol. 2005; 25:10684–94.46. Kim YI, Lee FN, Choi WS, Lee S, Youn JH. Insulin regulation of skeletal muscle PDK4 mRNA expression is impaired in acute insulin-resistant states. Diabetes. 2006; 55:2311–7.47. Zhang D, Li Y, Yao X, Wang H, Zhao L, Jiang H, et al. miR-182 regulates metabolic homeostasis by modulating glucose utilization in muscle. Cell Rep. 2016; 16:757–68.48. Wu P, Inskeep K, Bowker-Kinley MM, Popov KM, Harris RA. Mechanism responsible for inactivation of skeletal muscle pyruvate dehydrogenase complex in starvation and diabetes. Diabetes. 1999; 48:1593–9.49. Hwang B, Jeoung NH, Harris RA. Pyruvate dehydrogenase kinase isoenzyme 4 (PDHK4) deficiency attenuates the longterm negative effects of a high-saturated fat diet. Biochem J. 2009; 423:243–52.50. Jeoung NH, Harris RA. Pyruvate dehydrogenase kinase-4 deficiency lowers blood glucose and improves glucose tolerance in diet-induced obese mice. Am J Physiol Endocrinol Metab. 2008; 295:E46–54.51. Shi G, McQuibban GA. The mitochondrial rhomboid protease PARL is regulated by PDK2 to integrate mitochondrial quality control and metabolism. Cell Rep. 2017; 18:1458–72.52. Lee SJ, Jeong JY, Oh CJ, Park S, Kim JY, Kim HJ, et al. Pyruvate dehydrogenase kinase 4 promotes vascular calcification via SMAD1/5/8 phosphorylation. Sci Rep. 2015; 5:16577.53. Liu Z, Chen X, Wang Y, Peng H, Wang Y, Jing Y, et al. PDK4 protein promotes tumorigenesis through activation of cAMP-response element-binding protein (CREB)-Ras homolog enriched in brain (RHEB)-mTORC1 signaling cascade. J Biol Chem. 2014; 289:29739–49.54. Wu J, Zhao Y, Park YK, Lee JY, Gao L, Zhao J, et al. Loss of PDK4 switches the hepatic NF-κB/TNF pathway from prosurvival to pro-apoptosis. Hepatology. 2018; 68:1111–24.55. Connaughton S, Chowdhury F, Attia RR, Song S, Zhang Y, Elam MB, et al. Regulation of pyruvate dehydrogenase kinase isoform 4 (PDK4) gene expression by glucocorticoids and insulin. Mol Cell Endocrinol. 2010; 315:159–67.56. Crossland H, Constantin-Teodosiu D, Gardiner SM, Greenhaff PL. Peroxisome proliferator-activated receptor γ agonism attenuates endotoxaemia-induced muscle protein loss and lactate accumulation in rats. Clin Sci (Lond). 2017; 131:1437–47.57. Attaix D, Combaret L, Bechet D, Taillandier D. Role of the ubiquitin-proteasome pathway in muscle atrophy in cachexia. Curr Opin Support Palliat Care. 2008; 2:262–6.58. Hartmann-Petersen R, Gordon C. Proteins interacting with the 26S proteasome. Cell Mol Life Sci. 2004; 61:1589–95.59. Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001; 294:1704–8.60. Clavel S, Coldefy AS, Kurkdjian E, Salles J, Margaritis I, Derijard B. Atrophy-related ubiquitin ligases, atrogin-1 and MuRF1 are up-regulated in aged rat tibialis anterior muscle. Mech Ageing Dev. 2006; 127:794–801.61. Eddins MJ, Marblestone JG, Suresh Kumar KG, Leach CA, Sterner DE, Mattern MR, et al. Targeting the ubiquitin E3 ligase MuRF1 to inhibit muscle atrophy. Cell Biochem Biophys. 2011; 60:113–8.62. Sinam IS, Chanda D, Thoudam T, Kim MJ, Kim BG, Kang HJ, et al. Pyruvate dehydrogenase kinase 4 promotes ubiquitinproteasome system-dependent muscle atrophy. J Cachexia Sarcopenia Muscle. 2022; 13:3122–36.63. Kanda F, Okuda S, Matsushita T, Takatani K, Kimura KI, Chihara K. Steroid myopathy: pathogenesis and effects of growth hormone and insulin-like growth factor-I administration. Horm Res. 2001; 56 Suppl 1:24–8.64. Reincke M. Cushing syndrome associated myopathy: it is time for a change. Endocrinol Metab (Seoul). 2021; 36:564–71.65. Shen S, Liao Q, Liu J, Pan R, Lee SM, Lin L. Myricanol rescues dexamethasone-induced muscle dysfunction via a sirtuin 1-dependent mechanism. J Cachexia Sarcopenia Muscle. 2019; 10:429–44.66. Chen HH, Tsai LK, Liao KY, Wu TC, Huang YH, Huang YC, et al. Muscle-restricted nuclear receptor interaction protein knockout causes motor neuron degeneration through downregulation of myogenin at the neuromuscular junction. J Cachexia Sarcopenia Muscle. 2018; 9:771–85.67. Shi H, Verma M, Zhang L, Dong C, Flavell RA, Bennett AM. Improved regenerative myogenesis and muscular dystrophy in mice lacking Mkp5. J Clin Invest. 2013; 123:2064–77.68. Sacco A, Doyonnas R, Kraft P, Vitorovic S, Blau HM. Self-renewal and expansion of single transplanted muscle stem cells. Nature. 2008; 456:502–6.69. Adhikari A, Kim W, Davie J. Myogenin is required for assembly of the transcription machinery on muscle genes during skeletal muscle differentiation. PLoS One. 2021; 16:e0245618.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Anesthetic experience in a pediatric patient with pyruvate dehydrogenase complex (PDHC) deficiency: A case report
- The Role of Pyruvate Dehydrogenase Kinase in Diabetes and Obesity
- Role of the Pyruvate Dehydrogenase Complex in Metabolic Remodeling: Differential Pyruvate Dehydrogenase Complex Functions in Metabolism
- Inhibition of Sarcoplasmic Reticulum Ca2+ Uptake by Pyruvate and Fatty Acid in H9c2 Cardiomyocytes: Implications for Diabetic Cardiomyopathy
- Pyruvate Dehydrogenase Kinases: Therapeutic Targets for Diabetes and Cancers
