Lobeglitazone, A Peroxisome Proliferator-Activated Receptor-Gamma Agonist, Inhibits Papillary Thyroid Cancer Cell Migration and Invasion by Suppressing p38 MAPK Signaling Pathway
- Affiliations
-
- 1Division of Endocrinology and Metabolism, Department of Internal Medicine, Seoul St. Mary’s Hospital, College of Medicine, The Catholic University of Korea, Seoul, Korea
- KMID: 2521957
- DOI: http://doi.org/10.3803/EnM.2021.1155
Abstract
- Background
Peroxisome proliferator-activated receptor-gamma (PPAR-γ) ligands have been widely shown to correlate with epithelial-mesenchymal transition (EMT) and cancer progression. Lobeglitazone (LGZ) is a novel ligand of PPAR-γ; and its role in EMT and metastasis in papillary thyroid carcinoma (PTC) is poorly understood. We aimed to investigate the role of LGZ in metastatic behavior of PTC cells.
Methods
Half maximal inhibitory concentration (IC50) values of LGZ in BRAF-mutated PTC cell lines (BCPAP and K1) were determined using MTT assay. Rosiglitazone (RGZ), the PPAR-γ ligand was used as a positive control. The protein expression of PPAR-γ, cell-surface proteins (E-cadherin, N-cadherin), cytoskeletal protein (Vimentin), transcription factor (Snail), p38 mitogenactivated protein kinase (MAPK), extracellular signal-regulated kinase (ERK) 1/2 pathway, and matrix metalloproteinase (MMP)-2 expression were measured using Western blotting. Changes in E-cadherin expression were also determined using immunocytochemistry. Cell migration and invasion were analyzed using wound healing and Matrigel invasion assays.
Results
Treatment with LGZ or RGZ significantly inhibited transforming growth factor-beta1 (TGF-β1)-induced EMT-associated processes such as fibroblast-like morphological changes, EMT-related protein expression, and increased cell migration and invasion in BCPAP and K1 cells. LGZ restored TGF-β1-induced loss of E-cadherin, as observed using immunocytochemistry. Furthermore, LGZ and RGZ suppressed TGF-β1-induced MMP-2 expression and phosphorylation of p38 MAPK, but not ERK1/2. Although there was no change in PPAR-γ expression after treatment with LGZ or RGZ, the effect of downstream processes mediated by LGZ was hampered by GW9662, a PPAR-γ antagonist.
Conclusion
LGZ inhibits TGF-β1-induced EMT, migration, and invasion through the p38 MAPK signaling pathway in a PPAR-γ-dependent manner in PTC cells.
Keyword
Figure
-
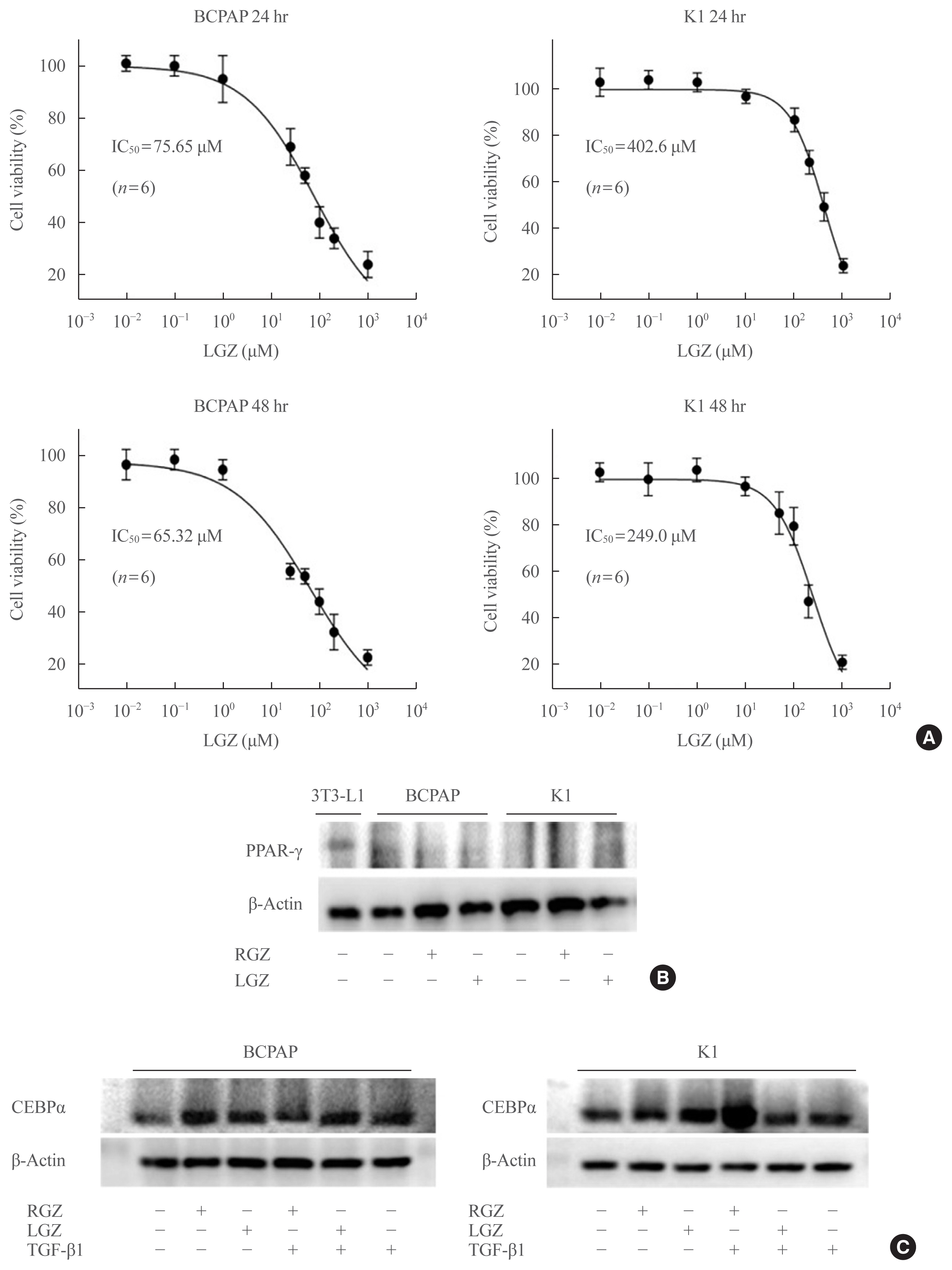
Fig. 1 Identification of lobeglitazone (LGZ) as a selective peroxisome proliferator-activated receptor (PPAR) agonist. (A) The half maximal inhibitory concentration (IC50) values of LGZ in papillary thyroid carcinoma (PTC) cells at 24 and 48 hours were measured using MTT assay. Data error bars correspond to the standard error of the mean (n=6). (B) Western blot assay shows the low protein level of PPAR-γ and no significant change in response to rosiglitazone (RGZ) or LGZ treatment, compared with control. BCPAP and K1 cells are treated with RGZ 10 μM or LGZ 10 μM for 24 hours. 3T3-L1 cell line is used as a positive control. (C) Western blot shows the protein levels of CCAAT/enhancer binding protein alpha (C/EBPα). Stimulation with transforming growth factor-beta1 (TGF-β1; 20 ng/mL) in the presence or absence of RGZ (10 μM) or LGZ (10 μM) at 24 hours in BCPAP and K1 cells.

Fig. 2 Transforming growth factor-beta1 (TGF-β1) induces epithelial-mesenchymal transition (EMT) in papillary thyroid carcinoma (PTC) cells. (A) BCPAP cells are treated with different concentrations of TGF-β1 for 48 hours and (B) K1 cells for 24 hours. Western blot analysis of E-cadherin, N-cadherin, Vimentin, and Snail protein expression in PTC cells. β-Actin is used as a control. Statistical analysis is performed using the Student’s t test from three independent experiments repeated in triplicate, and error bars correspond to the standard error of the mean and P values are indicated in each panel. aP<0.05; bP<0.01 vs. control.
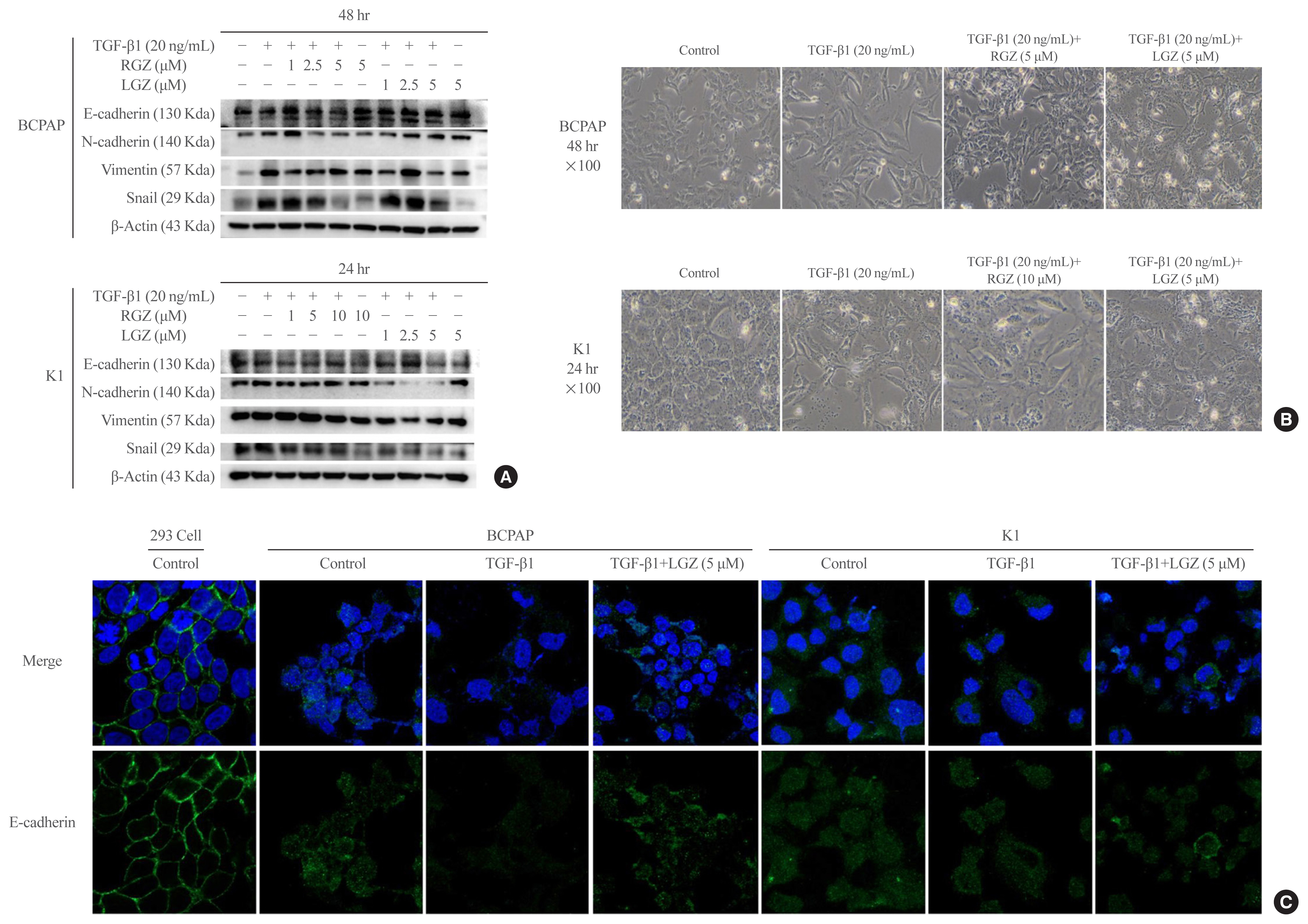
Fig. 3 Lobeglitazone (LGZ) and rosiglitazone (RGZ) inhibit transforming growth factor-beta1 (TGF-β1)-induced epithelial-mesenchymal transition in papillary thyroid carcinoma (PTC) cells. (A) Western blot analysis of E-cadherin, N-cadherin, Vimentin, Snail protein expression in PTC cells stimulated with TGF-β1 (20 ng/mL) in the presence or absence of peroxisome proliferator-activated receptor gamma ligands RGZ or LGZ at different concentrations for 24 and 48 hours. β-Actin is used as loading control. (B) The morphological changes in PTC cells following treatments at 24- and 48-hour time point. The images are acquired at 100× magnification. (C) Confocal microscopy images of E-cadherin (Alexa Fluor® 488, Green Fluorescent) expression under different stimuli in PTC cells. Human embryonic kidney 293 cells are used as positive control. Cell’s nuclei are stained with 4′,6-diamidino-2-phenylindole (DAPI). Data are expressed as mean±standard error of the mean for three experiments.
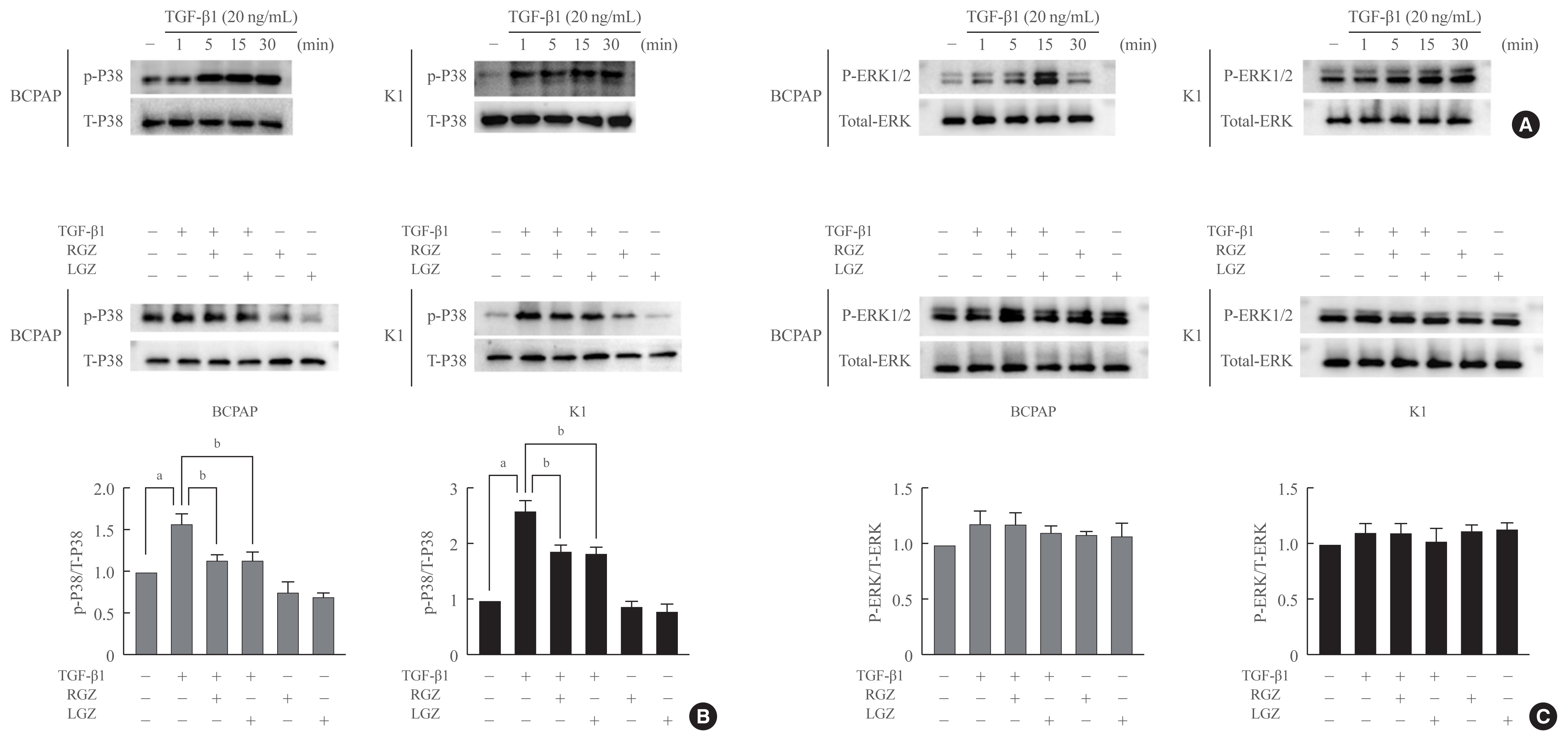
Fig. 4 Lobeglitazone (LGZ) and rosiglitazone (RGZ) inhibit transforming growth factor-beta1 (TGF-β1)-induced phosphorylation of p38 but not extracellular signal-regulated kinase (ERK) 1/2. (A) TGF-β1 induces activation of p38 and ERK1/2 in BCPAP and K1 cells in a time-dependent manner. (B) LGZ and RGZ inhibit TGF-β1-induced phosphorylation of p38. Cell are pretreated with RGZ or LGZ for 30 minutes and stimulated with or without TGF-β1 (20 ng/mL) for 15 minutes. Lower panel: relative protein expression rate was normalized using the value of p-P38/T-P38. (C) LGZ and RGZ have no suppressive effects on the TGF-β1-induced phosphorylation of ERK1/2. Cell are pretreated with RGZ or LGZ for 30 minutes and stimulated with or without TGF-β1 (20 ng/mL) for 15 minutes. Lower panel: relative protein expression rate was normalized using the value of p-ERK1/2/T-ERK. Statistical analysis is performed using the Student’s t test from three independent experiments repeated in triplicate; Error bars correspond to the standard error of the mean, P values are indicated in each panel. aP<0.05 vs. control; bP<0.05 vs. TGF-β1-treated.
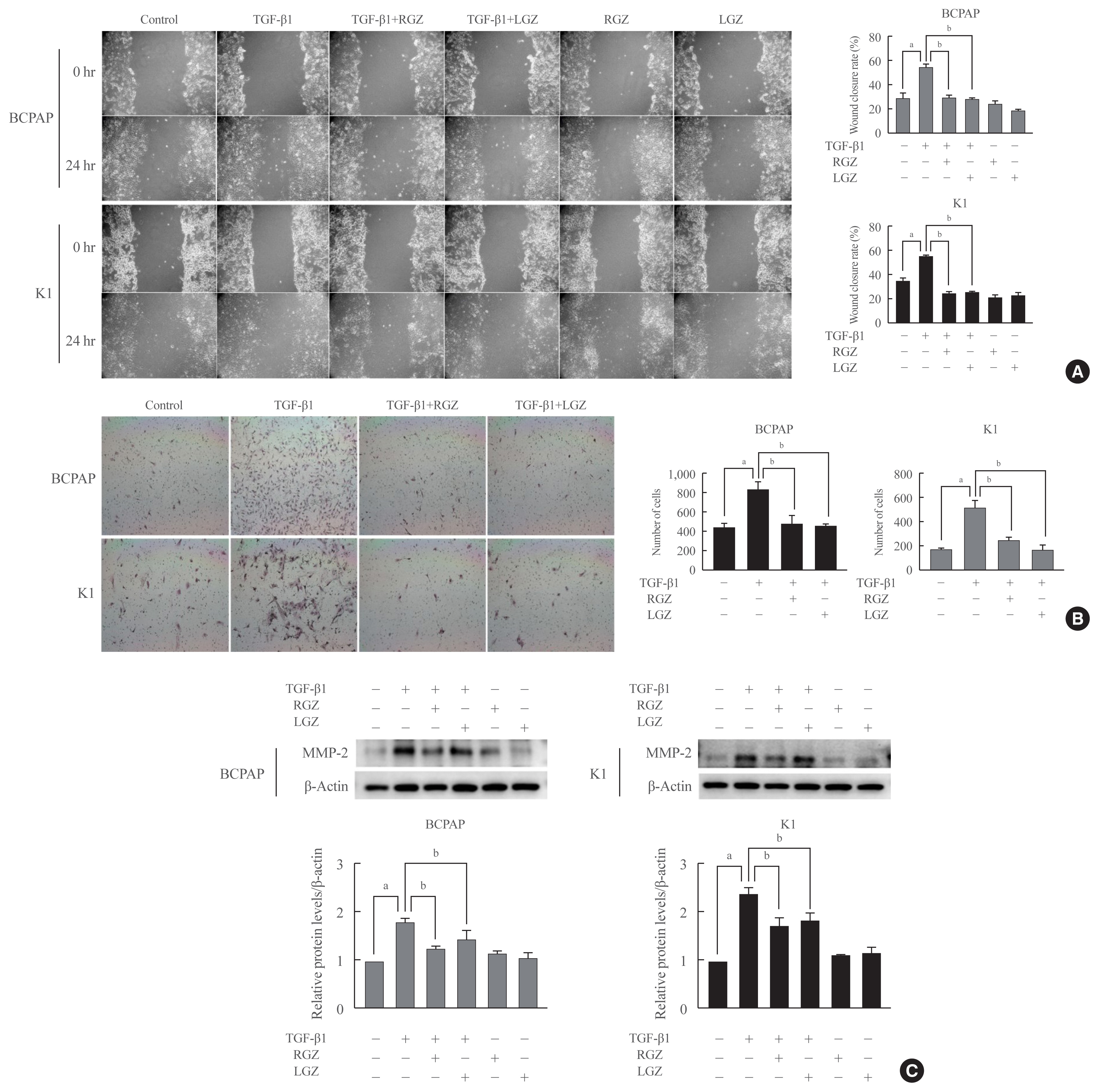
Fig. 5 Lobeglitazone (LGZ) and rosiglitazone (RGZ) inhibit cell migration, invasion, and matrix metalloproteinase 2 (MMP-2) protein expression induced by transforming growth factor-beta1 (TGF-β1) in papillary thyroid carcinoma (PTC) cells. (A) Wound healing cell migration assay shows that LGZ and RGZ inhibit PTC cell migration induced by TGF-β1. Left: representative images of scratched and recovering wounded areas at 0 and 24 hours. Right: quantitative analysis of the rate of wound closure. Cells treated with LGZ and RGZ show significantly lower migration ability than those treated with TGF-β1 (20 ng/mL). (B) Cells are trypsinized and plated in the Matrigel-coated transwell chambers to evaluate invasion. Left: representative images of PTC cells following different treatments at 24 hours. Right: quantitative analysis of the invasive cell number. Cells treated with peroxisome proliferator-activated receptor gamma (PPAR-γ) ligands show significantly lower invasion ability than those treated with TGF-β1. (C) Protein expression level of MMP-2 in PTC cells treated with TGF-β1 in the presence or absence of PPAR-γ ligands. Relative protein expression rate isnormalized using β-actin as a loading control. Data are expressed as mean±standard error of the mean for three experiments. Statistical analysis is performed using the Student’s t test, P values are indicated in each panel. aP<0.05 vs. control; bP<0.05 vs. TGF-β1-treated.

Fig. 6 Effects of lobeglitazone (LGZ) inhibition of phosphorylation of p38 mitogen-activated protein kinase (MAPK) and matrix metalloproteinase 2 (MMP-2) expression induced by transforming growth factor-beta1 (TGF-β1) are peroxisome proliferator-activated receptor gamma (PPAR-γ) dependent. (A) Western blot showing the effect of GW9662 on LGZ-mediated inhibition of the TGF-β1 activation of p38 MAPK and MMP-2 expression in papillary thyroid carcinoma (PTC) cells. PTC cells are pre-treated with GW9662 for 2 hours and cells are treated with presence or absence of LGZ 5 μM for 30 minutes, then stimulated with TGF-β1 (20 ng/mL) for 15 minutes. (B) The effect of GW9662 on LGZ-mediated inhibition of TGF-β1-induced MMP-2 protein expression in PTC cells by Western blot assay. PTC cells are pre-treated with GW9662 for 2 hours and cells are treated with presence or absence of LGZ 5 μM for 30 minutes, then stimulated with TGF-β1 at the indicated concentrations for 24 and 48 hours. Relative protein expression rate is normalized using β-actin as a loading control. Statistical analysis was performed using the Student’s t test from three independent experiments repeated in triplicates; Error bars correspond to the standard error of the mean; P values are indicated in each panel. aP<0.05 vs. control; bP<0.05 vs. TGF-β1-treated; cP<0.05 vs. TGF-β1 with LGZ-treated.
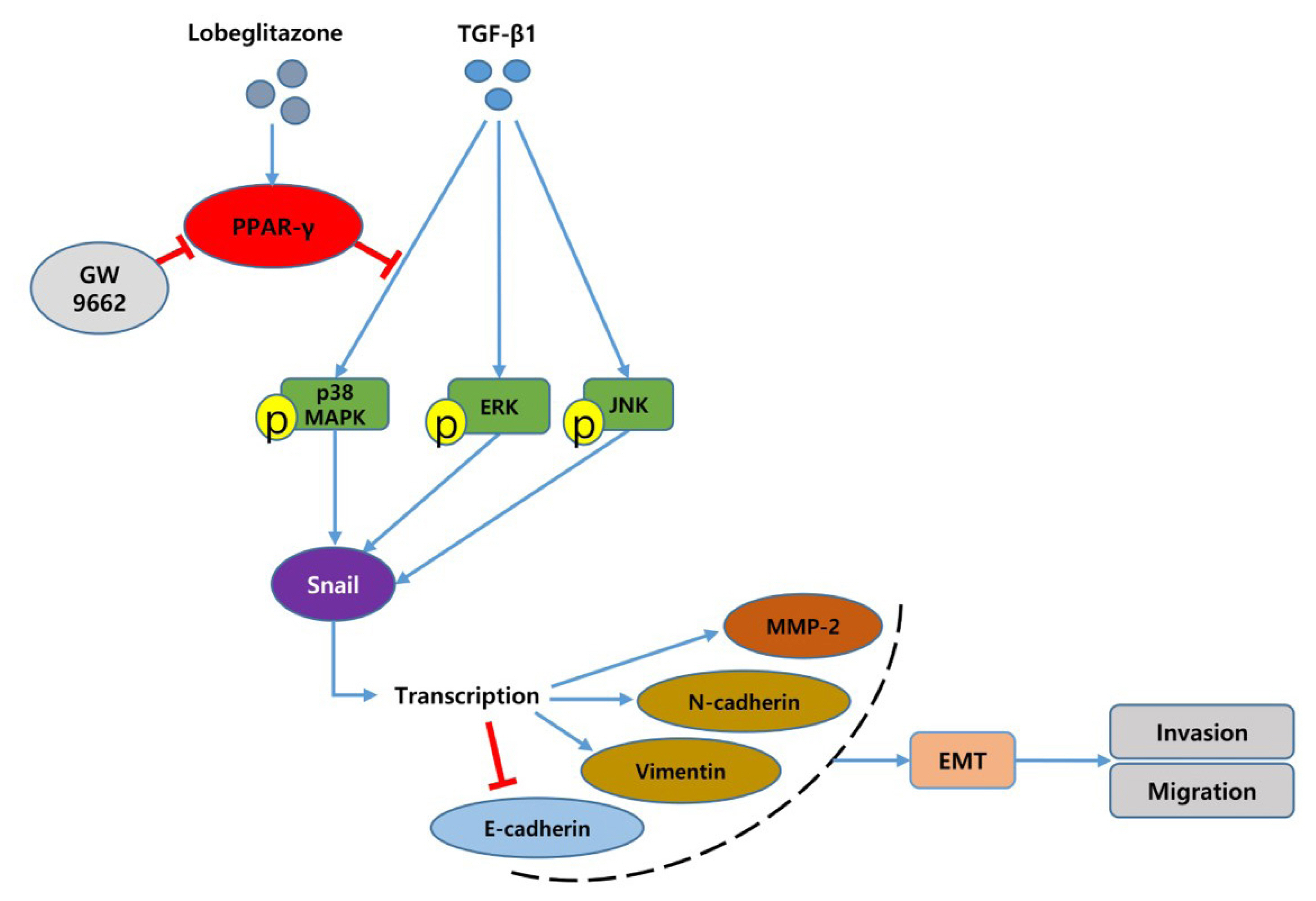
Fig. 7 Schematic representation of transforming growth factor-beta1 (TGF-β1)-induced epithelial-mesenchymal transition (EMT) and role of lobeglitazone (LGZ), a peroxisome proliferator-activated receptor gamma (PPAR-γ) agonist in BRAFV600E-mutated papillary thyroid carcinoma cells. LGZ suppresses the p38 mitogen-activated protein kinase (MAPK) signaling pathway and finally reduces migration and invasion of papillary thyroid carcinoma (PTC) cells induced by TGF-β1. “→” indicates promotion; “⊥” indicates inhibition. ERK, extracellular signal-regulated kinase.
Reference
-
1. Pellegriti G, Frasca F, Regalbuto C, Squatrito S, Vigneri R. Worldwide increasing incidence of thyroid cancer: update on epidemiology and risk factors. J Cancer Epidemiol. 2013; 2013:965212.
Article2. Lubitz CC, Sosa JA. The changing landscape of papillary thyroid cancer: epidemiology, management, and the implications for patients. Cancer. 2016; 122:3754–9.
Article3. Morris LG, Tuttle RM, Davies L. Changing trends in the incidence of thyroid cancer in the United States. JAMA Otolaryngol Head Neck Surg. 2016; 142:709–11.
Article4. LiVolsi VA. Papillary thyroid carcinoma: an update. Mod Pathol. 2011; 24(Suppl 2):S1–9.
Article5. Lebastchi AH, Callender GG. Thyroid cancer. Curr Probl Cancer. 2014; 38:48–74.
Article6. Xu J, Lamouille S, Derynck R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009; 19:156–72.7. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009; 139:871–90.
Article8. Wei SC, Fattet L, Yang J. The forces behind EMT and tumor metastasis. Cell Cycle. 2015; 14:2387–8.
Article9. Vu T, Datta PK. Regulation of EMT in colorectal cancer: a culprit in metastasis. Cancers (Basel). 2017; 9:171.
Article10. Javelaud D, Mauviel A. Mammalian transforming growth factor-betas: Smad signaling and physio-pathological roles. Int J Biochem Cell Biol. 2004; 36:1161–5.11. Vasko V, Espinosa AV, Scouten W, He H, Auer H, Liyanarachchi S, et al. Gene expression and functional evidence of epithelial-to-mesenchymal transition in papillary thyroid carcinoma invasion. Proc Natl Acad Sci U S A. 2007; 104:2803–8.
Article12. Riesco-Eizaguirre G, Rodriguez I, De la Vieja A, Costamagna E, Carrasco N, Nistal M, et al. The BRAFV600E oncogene induces transforming growth factor beta secretion leading to sodium iodide symporter repression and increased malignancy in thyroid cancer. Cancer Res. 2009; 69:8317–25.13. Knauf JA, Sartor MA, Medvedovic M, Lundsmith E, Ryder M, Salzano M, et al. Progression of BRAF-induced thyroid cancer is associated with epithelial-mesenchymal transition requiring concomitant MAP kinase and TGFβ signaling. Oncogene. 2011; 30:3153–62.
Article14. Atfi A, Djelloul S, Chastre E, Davis R, Gespach C. Evidence for a role of Rho-like GTPases and stress-activated protein kinase/c-Jun N-terminal kinase (SAPK/JNK) in transforming growth factor beta-mediated signaling. J Biol Chem. 1997; 272:1429–32.15. Fu H, He Y, Qi L, Chen L, Luo Y, Chen L, et al. cPLA2α activates PI3K/AKT and inhibits Smad2/3 during epithelial-mesenchymal transition of hepatocellular carcinoma cells. Cancer Lett. 2017; 403:260–70.
Article16. Werb Z. ECM and cell surface proteolysis: regulating cellular ecology. Cell. 1997; 91:439–42.
Article17. Aschebrook-Kilfoy B, Sabra MM, Brenner A, Moore SC, Ron E, Schatzkin A, et al. Diabetes and thyroid cancer risk in the National Institutes of Health-AARP Diet and Health Study. Thyroid. 2011; 21:957–63.
Article18. Hsu IR, Kim SP, Kabir M, Bergman RN. Metabolic syndrome, hyperinsulinemia, and cancer. Am J Clin Nutr. 2007; 86:s867–71.
Article19. Tseng CH. Rosiglitazone may reduce thyroid cancer risk in patients with type 2 diabetes. Ann Med. 2013; 45:539–44.
Article20. Grommes C, Landreth GE, Heneka MT. Antineoplastic effects of peroxisome proliferator-activated receptor gamma agonists. Lancet Oncol. 2004; 5:419–29.21. Reka AK, Kurapati H, Narala VR, Bommer G, Chen J, Standiford TJ, et al. Peroxisome proliferator-activated receptor-gamma activation inhibits tumor metastasis by antagonizing Smad3-mediated epithelial-mesenchymal transition. Mol Cancer Ther. 2010; 9:3221–32.22. Elstner E, Muller C, Koshizuka K, Williamson EA, Park D, Asou H, et al. Ligands for peroxisome proliferator-activated receptorgamma and retinoic acid receptor inhibit growth and induce apoptosis of human breast cancer cells in vitro and in BNX mice. Proc Natl Acad Sci U S A. 1998; 95:8806–11.23. Giordano TJ, Haugen BR, Sherman SI, Shah MH, Caoili EM, Koenig RJ. Pioglitazone therapy of PAX8-PPARγ fusion protein thyroid carcinoma. J Clin Endocrinol Metab. 2018; 103:1277–81.
Article24. Kato Y, Ying H, Zhao L, Furuya F, Araki O, Willingham MC, et al. PPARgamma insufficiency promotes follicular thyroid carcinogenesis via activation of the nuclear factor-kappaB signaling pathway. Oncogene. 2006; 25:2736–47.
Article25. Aiello A, Pandini G, Frasca F, Conte E, Murabito A, Sacco A, et al. Peroxisomal proliferator-activated receptor-gamma agonists induce partial reversion of epithelial-mesenchymal transition in anaplastic thyroid cancer cells. Endocrinology. 2006; 147:4463–75.26. Lee JH, Noh CK, Yim CS, Jeong YS, Ahn SH, Lee W, et al. Kinetics of the absorption, distribution, metabolism, and excretion of lobeglitazone, a novel activator of peroxisome proliferator-activated receptor gamma in rats. J Pharm Sci. 2015; 104:3049–59.
Article27. Moon KS, Lee JE, Lee HS, Hwang IC, Kim DH, Park HK, et al. CKD-501, a novel selective PPARγ agonist, shows no carcinogenic potential in ICR mice following oral administration for 104 weeks. J Appl Toxicol. 2014; 34:1271–84.
Article28. Lee HS, Chang M, Lee JE, Kim W, Hwang IC, Kim DH, et al. Carcinogenicity study of CKD-501, a novel dual peroxisome proliferator-activated receptors α and γ agonist, following oral administration to Sprague Dawley rats for 94–101 weeks. Regul Toxicol Pharmacol. 2014; 69:207–16.29. Nemenoff RA. Peroxisome proliferator-activated receptor-gamma in lung cancer: defining specific versus “off-target” effectors. J Thorac Oncol. 2007; 2:989–92.30. Han S, Roman J. Rosiglitazone suppresses human lung carcinoma cell growth through PPARgamma-dependent and PPARgamma-independent signal pathways. Mol Cancer Ther. 2006; 5:430–7.31. Bae KH, Seo JB, Jung YA, Seo HY, Kang SH, Jeon HJ, et al. Lobeglitazone, a novel peroxisome proliferator-activated receptor γ agonist, attenuates renal fibrosis caused by unilateral ureteral obstruction in mice. Endocrinol Metab (Seoul). 2017; 32:115–23.
Article32. Vella V, Nicolosi ML, Giuliano S, Bellomo M, Belfiore A, Malaguarnera R. PPAR-γ agonists as antineoplastic agents in cancers with dysregulated IGF axis. Front Endocrinol (Lausanne). 2017; 8:31.
Article33. Feinstein DL, Spagnolo A, Akar C, Weinberg G, Murphy P, Gavrilyuk V, et al. Receptor-independent actions of PPAR thiazolidinedione agonists: is mitochondrial function the key? Biochem Pharmacol. 2005; 70:177–88.
Article34. Wood WM, Sharma V, Bauerle KT, Pike LA, Zhou Q, Fretwell DL, et al. PPARγ promotes growth and invasion of thyroid cancer cells. PPAR Res. 2011; 2011:171765.35. Tontonoz P, Hu E, Spiegelman BM. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell. 1994; 79:1147–56.
Article36. Sugii S, Olson P, Sears DD, Saberi M, Atkins AR, Barish GD, et al. PPARgamma activation in adipocytes is sufficient for systemic insulin sensitization. Proc Natl Acad Sci U S A. 2009; 106:22504–9.37. Lefterova MI, Zhang Y, Steger DJ, Schupp M, Schug J, Cristancho A, et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008; 22:2941–52.38. Miller M, Shuman JD, Sebastian T, Dauter Z, Johnson PF. Structural basis for DNA recognition by the basic region leucine zipper transcription factor CCAAT/enhancer-binding protein alpha. J Biol Chem. 2003; 278:15178–84.39. Dobson ME, Diallo-Krou E, Grachtchouk V, Yu J, Colby LA, Wilkinson JE, et al. Pioglitazone induces a proadipogenic antitumor response in mice with PAX8-PPARgamma fusion protein thyroid carcinoma. Endocrinology. 2011; 152:4455–65.
Article40. Sato A, Yamada N, Ogawa Y, Ikegami M. CCAAT/enhancer-binding protein-α suppresses lung tumor development in mice through the p38α MAP kinase pathway. PLoS One. 2013; 8:e57013.
Article41. Lu J, Du C, Yao J, Wu B, Duan Y, Zhou L, et al. C/EBPα suppresses lung adenocarcinoma cell invasion and migration by inhibiting β-catenin. Cell Physiol Biochem. 2017; 42:1779–88.
Article42. Saiselet M, Floor S, Tarabichi M, Dom G, Hebrant A, van Staveren WC, et al. Thyroid cancer cell lines: an overview. Front Endocrinol (Lausanne). 2012; 3:133.
Article43. Lourenco AR, Roukens MG, Seinstra D, Frederiks CL, Pals CE, Vervoort SJ, et al. C/EBPα is crucial determinant of epithelial maintenance by preventing epithelial-to-mesenchymal transition. Nat Commun. 2020; 11:785.
Article44. Qi C, Zhu Y, Reddy JK. Peroxisome proliferator-activated receptors, coactivators, and downstream targets. Cell Biochem Biophys. 2000; 32:187–204.
Article45. Onder TT, Gupta PB, Mani SA, Yang J, Lander ES, Weinberg RA. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008; 68:3645–54.
Article46. Lee HJ, Su Y, Yin PH, Lee HC, Chi CW. PPAR(gamma)/PGC-1(alpha) pathway in E-cadherin expression and motility of HepG2 cells. Anticancer Res. 2009; 29:5057–63.47. Chang SN, Lee JM, Oh H, Kim U, Ryu B, Park JH. Troglitazone inhibits the migration and invasion of PC-3 human prostate cancer cells by upregulating E-cadherin and glutathione peroxidase 3. Oncol Lett. 2018; 16:5482–8.
Article48. Zhou Q, Chen J, Feng J, Xu Y, Zheng W, Wang J. SOSTDC1 inhibits follicular thyroid cancer cell proliferation, migration, and EMT via suppressing PI3K/Akt and MAPK/Erk signaling pathways. Mol Cell Biochem. 2017; 435:87–95.
Article49. Tse JC, Kalluri R. Mechanisms of metastasis: epithelial-to-mesenchymal transition and contribution of tumor microenvironment. J Cell Biochem. 2007; 101:816–29.
Article50. Palona I, Namba H, Mitsutake N, Starenki D, Podtcheko A, Sedliarou I, et al. BRAFV600E promotes invasiveness of thyroid cancer cells through nuclear factor kappaB activation. Endocrinology. 2006; 147:5699–707.
Article51. Zou M, Baitei EY, BinEssa HA, Al-Mohanna FA, Parhar RS, St-Arnaud R, et al. Cyp24a1 attenuation limits progression of BrafV600E-induced papillary thyroid cancer cells and sensitizes them to BRAFV600E inhibitor PLX4720. Cancer Res. 2017; 77:2161–72.52. Jorda M, Olmeda D, Vinyals A, Valero E, Cubillo E, Llorens A, et al. Upregulation of MMP-9 in MDCK epithelial cell line in response to expression of the Snail transcription factor. J Cell Sci. 2005; 118(Pt 15):3371–85.
Article53. Xie L, Law BK, Chytil AM, Brown KA, Aakre ME, Moses HL. Activation of the Erk pathway is required for TGF-beta1-induced EMT in vitro. Neoplasia. 2004; 6:603–10.
Article54. Yu L, Hebert MC, Zhang YE. TGF-beta receptor-activated p38 MAP kinase mediates Smad-independent TGF-beta responses. EMBO J. 2002; 21:3749–59.
Article55. Kumar B, Koul S, Petersen J, Khandrika L, Hwa JS, Meacham RB, et al. p38 mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity. Cancer Res. 2010; 70:832–41.
Article56. Chapnick DA, Warner L, Bernet J, Rao T, Liu X. Partners in crime: the TGFβ and MAPK pathways in cancer progression. Cell Biosci. 2011; 1:42.
Article57. Ciaramella V, Sasso FC, Di Liello R, Corte CM, Barra G, Viscardi G, et al. Activity and molecular targets of pioglitazone via blockade of proliferation, invasiveness and bioenergetics in human NSCLC. J Exp Clin Cancer Res. 2019; 38:178.
Article58. Nuwormegbe SA, Sohn JH, Kim SW. A PPAR-gamma agonist rosiglitazone suppresses fibrotic response in human pterygium fibroblasts by modulating the p38 MAPK pathway. Invest Ophthalmol Vis Sci. 2017; 58:5217–26.
Article59. Poleni PE, Bianchi A, Etienne S, Koufany M, Sebillaud S, Netter P, et al. Agonists of peroxisome proliferators-activated receptors (PPAR) alpha, beta/delta or gamma reduce transforming growth factor (TGF)-beta-induced proteoglycans’ production in chondrocytes. Osteoarthritis Cartilage. 2007; 15:493–505.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Caspase Recruitment Domain Containing Protein 9 Suppresses Non-Small Cell Lung Cancer Proliferation and Invasion via Inhibiting MAPK/p38 Pathway
- Expression of Down Stream Molecules of RET (p-ERK, p-p38 MAPK, p-JNK and p-AKT) in Papillary Thyroid Carcinomas
- Role of Pexoxisome Proliferator Activated Receptor Gamma in Growth Regulation of Thyroid Cancer Cells
- Ras Mitogen-activated Protein Kinase Signaling and Kinase Suppressor of Ras as Therapeutic Targets for Hepatocellular Carcinoma
- Effect of Transforming Growth Factor (TGF)-beta and Peroxisome Proliferator-Activated Receptor (PPAR)-gamma in Endometrial Decidualization
