Disorders of Secondary Neurulation : Mainly Focused on Pathoembryogenesis
- Affiliations
-
- 1Neuro-oncology Clinic, Center for Rare Cancers, National Cancer Center, Goyang, Korea
- 2Division of Pediatric Neurosurgery, Seoul National University Children’s Hospital, Seoul, Korea
- 3Department of Anatomy, Seoul National University College of Medicine, Seoul, Korea
- KMID: 2515495
- DOI: http://doi.org/10.3340/jkns.2021.0023
Abstract
- Recent advancements in basic research on the process of secondary neurulation and increased clinical experience with caudal spinal anomalies with associated abnormalities in the surrounding and distal structures shed light on further understanding of the pathoembryogenesis of the lesions and led to the new classification of these dysraphic entities. We summarized the changing concepts of lesions developed from the disordered secondary neurulation shown during the last decade. In addition, we suggested our new pathoembryogenetic explanations for a few entities based on the literature and the data from our previous animal research. Disordered secondary neurulation at each phase of development may cause corresponding lesions, such as failed junction with the primary neural tube (junctional neural tube defect and segmental spinal dysgenesis), dysgenesis or duplication of the caudal cell mass associated with disturbed activity of caudal mesenchymal tissue (caudal agenesis and caudal duplication syndrome), failed ingression of the primitive streak to the caudal cell mass (myelomeningocele), focal limited dorsal neuro-cutaneous nondisjunction (limited dorsal myeloschisis and congenital dermal sinus), neuro-mesenchymal adhesion (lumbosacral lipomatous malformation), and regression failure spectrum of the medullary cord (thickened filum and filar cyst, low-lying conus, retained medullary cord, terminal myelocele and terminal myelocystocele). It seems that almost every anomalous entity of the primary neural tube may occur in the area of secondary neurulation. Furthermore, the close association with the activity of caudal mesenchymal tissue in secondary neurulation involves a wider range of surrounding structures than in primary neurulation. Although the majority of the data are from animals, not from humans and many theories are still conjectural, these changing concepts of normal and disordered secondary neurulation will provoke further advancements in our management strategies as well as in the pathoembryogenetic understanding of anomalous lesions in this area.
Figure
-
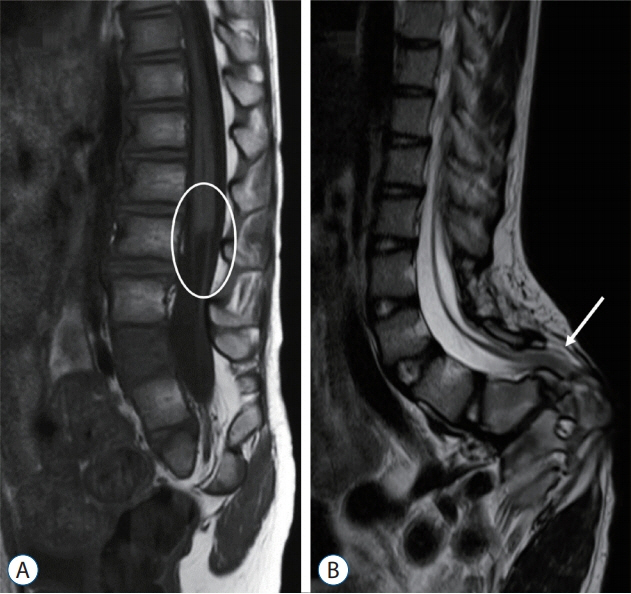
Fig. 1. A : A T1 sagittal MRI image showing a blunt-ended conus at the L1 level (circle). B : A T2 sagittal MRI image showing a fatty filum with a low-level of the conus (arrow). MRI : magnetic resonance imaging.

Fig. 2. Common manifestations of caudal duplication syndrome. The proximal portion of duplicated organs is separated by a septum but not as two different independent organs. In contrast to two urethral, vaginal and anal orifices (A), duplicated colons run in parallel, sharing one septum between them (B). Similarly, duplicated bladders or uteri share a septum in the midline separating the two (C). Duplication occurs similar to the branching of a tree, followed by the formation of two sets of suborgans, such as the urethra, vagina and anus, until reaching the distal end. Reprinted from Harris et al. [24] with permission from Sage publications, Inc.

Fig. 3. A schematic diagram of the hindgut including cloaca and medullary cord during the formation of the urogenital sinus and anorectal canal in the normal state (A) and in caudal duplication syndrome (CDupS) (B). The upper large circle indicates the cloaca, and the lower small circle(s) indicates the medullary cord. The arrow indicates the movement of mesenchymal cells, whereas the length of the arrow indicates the power of the movement. Arrows numbered ① indicate the formation of the septum (normally urorectal septum, additional midline septum in CDupS) in the cloaca by the mesenchymal push from both lateral walls in normal development and from the posterior wall as well in CDupS. Arrows numbered ② indicate the ventral push of the caudal mesenchyme. Duplicated medullary cords give rise to an aberrant insertion of mesenchymal tissue between the two medullary cords at the midline (B), which causes abnormal midline septation of the cloaca (or hindgut). Note that the ventral push of the mesenchyme around the lateral sides of the medullary cords in CDupS (B) is weaker than that in the normal state (A) due to the leakage of mesenchymal growth power through the midline. Weak ventral push along the lateral sides of the cloaca makes the urorectal septum locate more posteriorly than in the normal position, as in caudal agenesis, which may later result in an imperforate anus. Reprinted from Yang et al. [69] with permission from Springer Nature. MC : medullary cord.
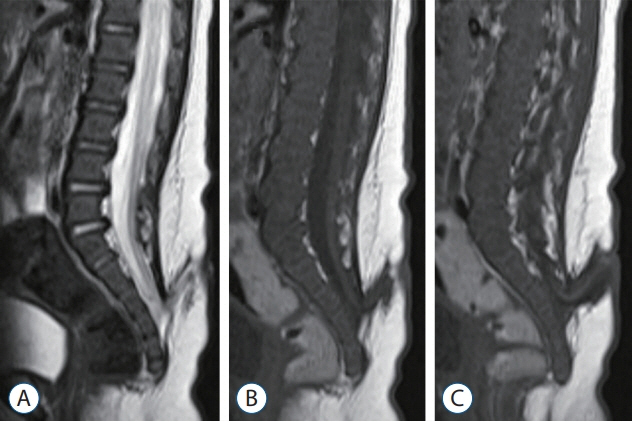
Fig. 4. A 2-month-old girl who was born with a sacral myelomeningocele (MMC) and underwent a delayed operation : T2 (A) and T1 (B and C) magnetic resonance imaging sections show a typical feature of an MMC, but the bony defect is low, below S3. The distal spinal cord turns to the cranial side.
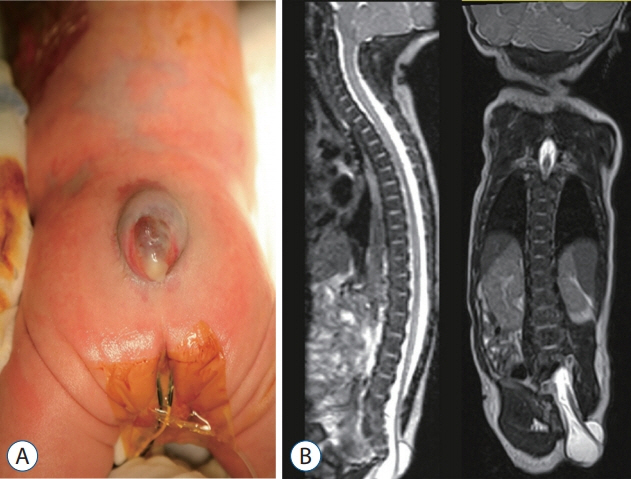
Fig. 5. A 1-day-old girl with a fetal diagnosis of a myelomeningocele. A : A gross photo showing the cystic sac with a skin defect at the sacrococcygeal level. B : T2 sagittal and coronal sections of magnetic resonance imaging showing the low-lying cord protruding through the fascia defect at the S2 level and below, going straight to the caudal side and attached to the skin defect area.
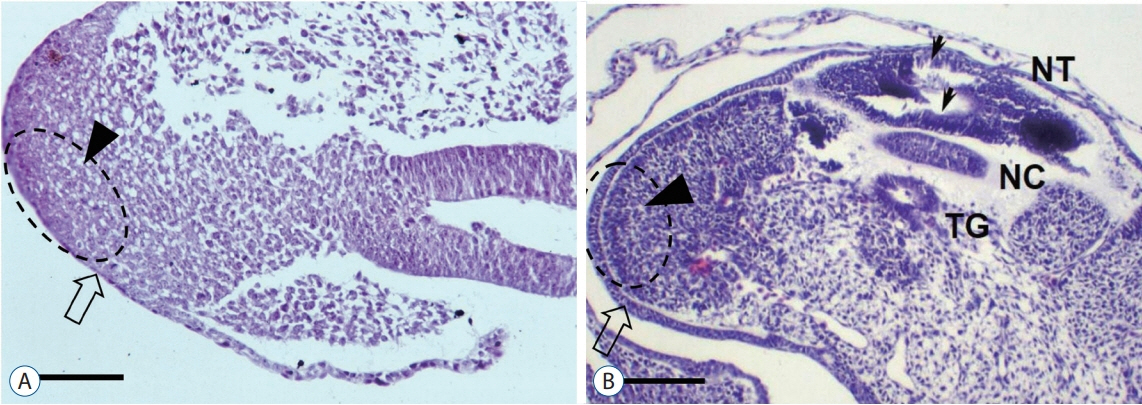
Fig. 6. The surface ectoderm (open arrow) and the caudal cell mass (arrowhead) are continuous in a Hamburger and Hamilton (H-H) stage 16 chick embryo (A; H&E, scale bar=100 μm). At H-H stage 20, the two layers are clearly divided (B; H&E, scale bar=100 μm). Small arrows indicate multiple vacuoles in the secondary neural tube. Reprinted from Kim et al. [27] by permission of the Congress of Neurological Surgeons. NT : neural tube, NC : notochord, TG : tailgut.
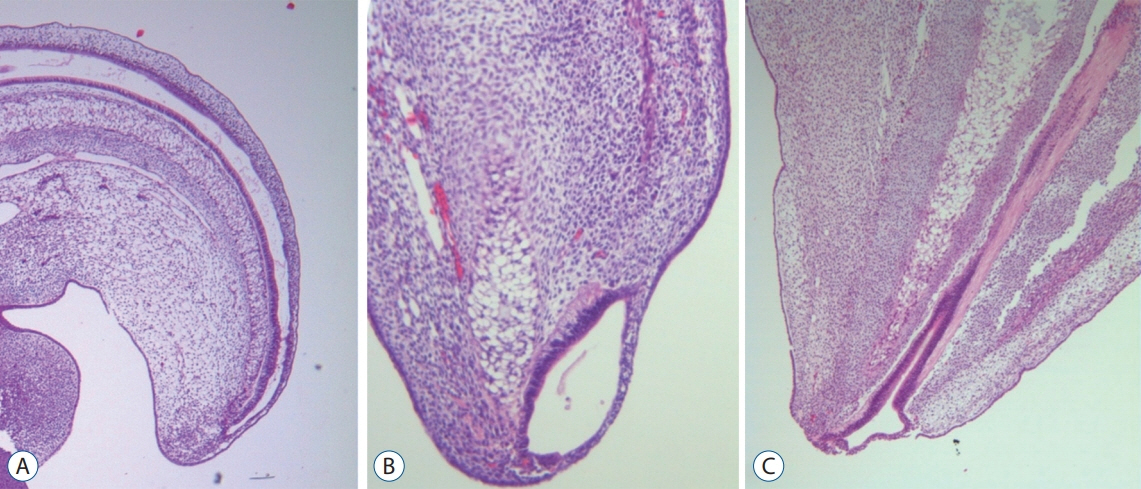
Fig. 7. Serial images of chick embryos showing the central canal of the medullary cord (A; H&E, ×40), a terminal balloon seen as a focal dilatation of the distal end of the central canal (B; H&E, ×200), and the shrinkage of the terminal balloon (C; H&E, ×200). Modified from Lee et al. [32] with reprint permission from Springer Nature.

Fig. 8. A schematic drawing showing the pathoembryogenesis of limited dorsal myeloschisis (LDM) compared with congenital dermal sinus (CDS). The common basic error is a failure of normal neurocutaneous disjunction during primary neurulation. If the neural tissue is pulled up to the skin side, it results in LDM (A), whereas if the cutaneous tissue is pulled down to the neural side, it leads to CDS (B).
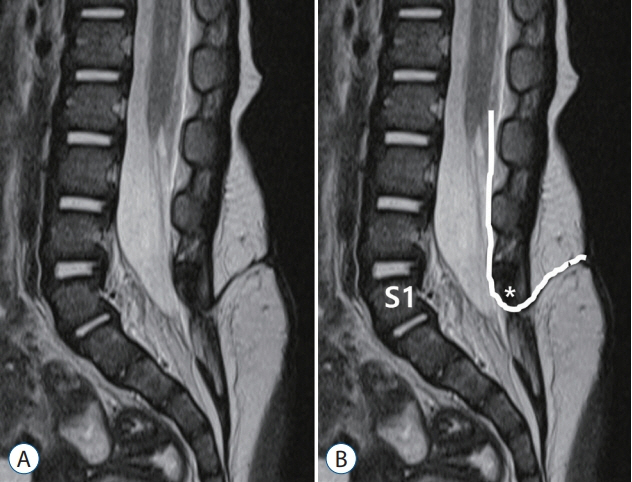
Fig. 9. A drawing showing the method to determine the spinal level of limited dorsal myeloschisis (LDM). A : T2 sagittal image. B : A method of counting the level of LDM. An LDM stalk traverses through the S1–2 interspinous ligament. The spinal level of this LDM was regarded as S1. Asterisk indicate dysplastic small spinous process of S1.

Fig. 10. Cases with low-lying limited dorsal myeloschisis (LDM). The spinal levels of both LDM cases are S3. The LDM stalks were attached to the low-lying coni. Green line indicate level of S1–2 junction (the LDM tracts passing though the interspinous spaces below this line seem attached to the spinal cord below S1–2 junction which was known to be formed by secondary neurulation only, not by primary neurulation).
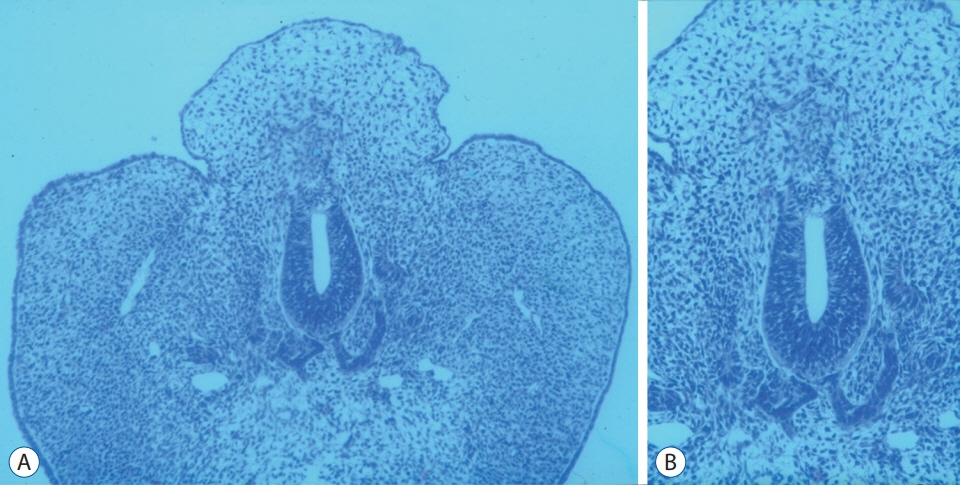
Fig. 11. A and B : A chick embryo showing histological findings suggestive of lumbosacral lipomatous malformation after the incision of the unilateral neural fold : a back hump making a sharp angle with surrounding normal skin, the blending of neuroepithelial and mesenchymal tissues through an indistinct basement membrane and an abnormal shape of the notochord (A : H&E, ×100; B : H&E, ×200). Reprinted from Li et al. [36] with permission from S. Karger AG. Basel.

Fig. 12. The junction between the primary neural tube and the CCM in a chick embryo of Hamburger and Hamilton stage 16 (a sagittal section, slightly off the midline). The CCM makes the medullary cord by the process of secondary neurulation from Hensen’s node to the caudal side of the embryo. H&E, scale bar=100 μm. 1 NT : primary neural tube, Jct Z : junctional zone between the primary and secondary neural tubes, CCM : caudal cell mass.

Fig. 13. A transverse section of a medullary cord in a chick embryo of Hamburger and Hamilton stage 30. The mesenchymal tissue between the medullary cord and skin is visible. There are still multiple vacuoles in the medullary cord. If mesenchymal tissue adheres to the dorsal aspect of the medullary cord during the process of secondary neurulation, lumbosacral lipomatous malformation of the transitional type will occur. H&E, scale bar=50 μm. Mes : mesenchymal tissue, MC : medullary cord.
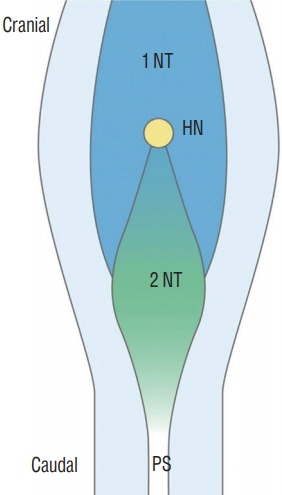
Fig. 14. A schematic drawing of Hensen’s node area in a chick embryo of Hamburger and Hamilton stage 8. The caudal part of Hensen’s node in the midline is the future secondary neural tube where the secondary neurulation progresses to the caudal side. The neural plates of the caudal primary neural tube are located at the lateral sides. If the caudal part of Hensen’s node (and the cranial part of the 'node-streak border' or ’rhomboidal area') has neuro-mesenchymal adhesion, the caudal end of primary neurulation is disturbed, and the fat is attached to the dorsal spinal cord up beyond the normal dorsocaudal end of the primary neural tube, which is known as the junction of S1–2 spinal cord segments. Modified from Shimokita and Takahashi [58] with permission from Japanese Society of Developmental Biologists. 1 NT : primary neural tube, HN : Hensen’s node, 2 NT : secondary neural tube, PS : primitive streak.

Fig. 15. Hamburger and Hamilton stage 30–35 chick embryos show the fully formed medullary cord with a terminal balloon (terminal vesicle) (A; H&E, scale bar=100 μm) and then regression of the balloon (B; H&E, scale bar=100 μm). TB : terminal balloon.

Fig. 16. A : A normal caudal spinal cord. B : If the site of neuro-mesenchymal adhesion at the caudal end of the medullary cord was pulled into the spinal canal through the sacral hiatus, a lumbosacral lipomatous malformation of the caudal type is formed. C : However, when abnormal neuro-mesenchymal adhesion occurs but there is no traction of mesenchymal tissue into the neural side, it may result in a typical retained medullary cord. D and E : If the spinal cord is distracted to the extraspinal side, a terminal myelocele (if the terminal balloon collapses, D) or terminal myelocystocele (if the terminal balloon persists, E) is formed.
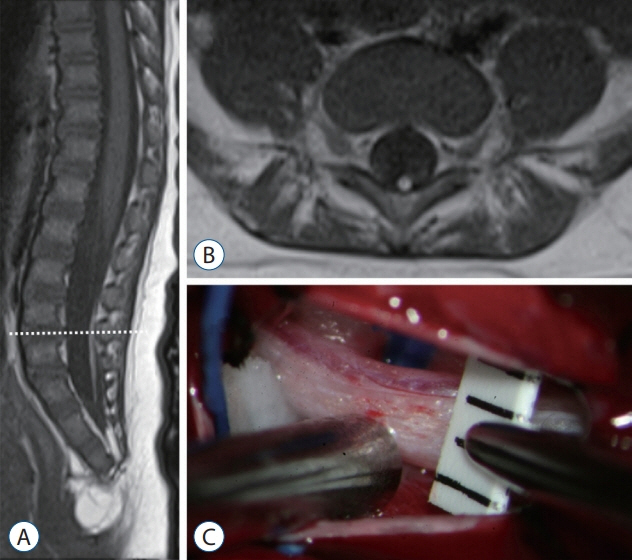
Fig. 17. A 3-month-old girl with filar lipoma. A : A T1 sagittal image shows the L2–3 level of conus medullaris tethered by the fatty filum. B : A T1 axial image at the level of the dotted line (A). C : An intraoperative photograph shows a fatty filum of 2.5 mm thickness.

Fig. 18. A 6-month old boy with elongated and right-side deviated gluteal folds. Sagittal (A) and axial (B) section sonographic images show a filar cyst immediately below the conus medullaris : a hypoechoic cyst (asterisk) in the filum 10.8 mm in length and 2.6 mm in diameter.
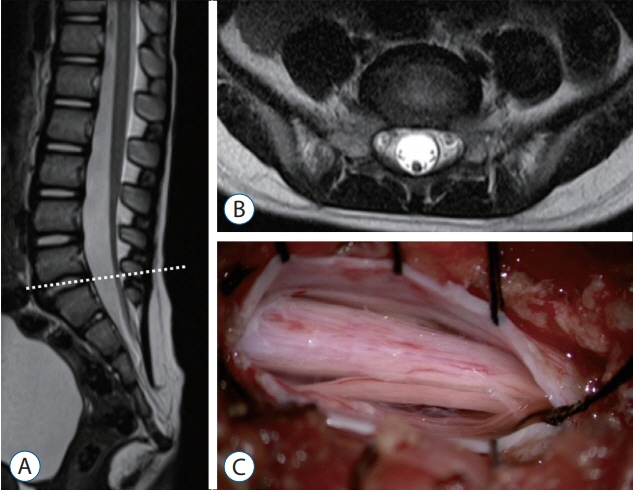
Fig. 19. Magnetic resonance imaging and an intraoperative photograph in a retained medullary cord patient. A : A T2 sagittal image. A cord-like structure extending to the sacral area. B : T2 axial images at the L5–S1 level (dotted line in A). C : Left L5 partial hemilaminectomy shows a thick, pia-covered medullary cord passing through. Reprinted from Kim et al. [28] with permission from the Korean Neurosurgical Society.

Fig. 20. Sagittal magnetic resonance imaging images of a case with terminal myelocele show the herniated spinal cord through laminar and fascial defects. Trumpet-like flaring is not seen. Only an enlarged subarachnoid space and herniated spinal cord and roots are observed. A : T2 sagittal image, midline plane. B : T2 sagittal image, paramedian plane where the herniated spinal cord is best seen. Reprinted from Lee et al. [32] by permission from Springer Nature.

Fig. 21. This (arrowheads) highlights the remarkable similarity between secondary neurulation in chick embryos (A; H&E, ×200) and the T2 sagittal image in a prototypical human terminal myelocystocele case (B). C : A T2 axial image. D : A T1 axial image.
Reference
-
References
1. Acer T, Ötgün İ, Sağnak Akıllı M, Gürbüz EE, Güney LH, Hiçsönmez A. A newborn with caudal duplication and duplex imperforate anus. J Pediatr Surg. 48:E37–E43. 2013.
Article2. Al-Omari MH, Eloqayli HM, Qudseih HM, Al-Shinag MK. Isolated lipoma of filum terminale in adults: MRI findings and clinical correlation. J Med Imaging Radiat Oncol. 55:286–290. 2011.
Article3. Al Alayet YF, Samujh R, Lyngdoh TS, Mansoor K, Al Kasim F, Al-Mustafa AA. An extremely rare case of classic complete caudal duplication: dipygus. J Indian Assoc Pediatr Surg. 19:169–171. 2014.
Article4. Arai H, Sato K, Okuda O, Miyajima M, Hishii M, Nakanishi H, et al. Surgical experience of 120 patients with lumbosacral lipomas. Acta Neurochir (Wien). 143:857–864. 2001.
Article5. Balioğlu MB, Akman YE, Ucpunar H, Albayrak A, Kargın D, Atıcı Y, et al. Sacral agenesis: evaluation of accompanying pathologies in 38 cases, with analysis of long-term outcomes. Childs Nerv Syst. 32:1693–1702. 2016.
Article6. Bannykh SI, Bannykh GI, Mannino FL, Jones KL, Hansen L, Benirschke K, et al. Partial caudal duplication in a newborn associated with meningomyelocele and complex heart anomaly. Teratology. 63:94–99. 2001.
Article7. Bansal G, Ghosh D, George U, Bhatti W. Unusual coexistence of caudal duplication and caudal regression syndromes. J Pediatr Surg. 46:256–258. 2011.
Article8. Bertocchini F, Stern CD. Gata2 provides an early anterior bias and uncovers a global positioning system for polarity in the amniote embryo. Development. 139:4232–4238. 2012.
Article9. Brown E, Matthes JC, Bazan C 3rd, Jinkins JR. Prevalence of incidental intraspinal lipoma of the lumbosacral spine as determined by MRI. Spine (Phila Pa 1976). 19:833–836. 1994.
Article10. Chapman PH. Congenital intraspinal lipomas. Pediatr Neurosurg. 9:37–47. 1982.
Article11. Copp AJ, Stanier P, Greene ND. Neural tube defects: recent advances, unsolved questions, and controversies. Lancet Neurol. 12:799–810. 2013.
Article12. Criley BB. Analysis of embryonic sources and mechanims of development of posterior levels of chick neural tubes. J Morphol. 128:465–501. 1969.
Article13. Dady A, Havis E, Escriou V, Catala M, Duband JL. Junctional neurulation: a unique developmental program shaping a discrete region of the spinal cord highly susceptible to neural tube defects. J Neurosci. 34:13208–13221. 2014.
Article14. Denton JR. The association of congenital spinal anomalies with imperforate anus. Clin Orthop Relat Res. (162):91–98. 1982.
Article15. Dominguez R, Rott J, Castillo M, Pittaluga RR, Corriere JN Jr. Caudal duplication syndrome. Am J Dis Child. 147:1048–1052. 1993.
Article16. Economides KD, Zeltser L, Capecchi MR. Hoxb13 mutations cause overgrowth of caudal spinal cordand tail vertebrae. Dev Biol. 256:317–330. 2003.
Article17. Eibach S, Moes G, Hou YJ, Zovickian J, Pang D. Unjoined primary and secondary neural tubes: junctional neural tube defect, a new form of spinal dysraphism caused by disturbance of junctional neurulation. Childs Nerv Syst. 33:1633–1647. 2017.
Article18. Emery JL, Lendon RG. Lipomas of the cauda equina and other fatty tumours related to neurospinal dysraphism. Dev Med Child Neurol Suppl. 20:62–70. 1969.
Article19. Finger T, Schaumann A, Grillet F, Schulz M, Thomale UW. Retethering after transection of a tight filum terminale, postoperative MRI may help to identify patients at risk. Childs Nerv Syst. 36:1499–1506. 2020.
Article20. Gould GM, Pyle WL. Prenatal anomalies. Anomalies and curiosities of medicine. New York: The Julian Press, Inc;1896.21. Griffith CM, Sanders EJ. Effects of extracellular matrix components on the differentiation of chick embryo tail bud mesenchyme in culture. Differentiation. 47:61–68. 1991.
Article22. Griffith CM, Wiley MJ. Direct effects of retinoic acid on the development of the tail bud in chick embryos. Teratology. 39:261–275. 1989.
Article23. Griffith CM, Wiley MJ, Sanders EJ. The vertebrate tail bud: three germ layers from one tissue. Anat Embryol (Berl). 185:101–113. 1992.
Article24. Harris J, Blackwood B, Pillai S, Kanard R. Caudal duplication: management of a rare congenital condition. Am Surg. 82:E227–E229. 2016.
Article25. Hughes AF, Freeman RB. Comparative remarks on the development of the tail cord among higher vertebrates. J Embryol Exp Morphol. 32:355–363. 1974.
Article26. Irani N, Goud AR, Lowe LH. Isolated filar cyst on lumbar spine sonography in infants: a case-control study. Pediatr Radiol. 36:1283–1288. 2006.
Article27. Kim JW, Wang KC, Chong S, Kim SK, Lee JY. Limited dorsal myeloschisis: reconsideration of its embryological origin. Neurosurgery. 86:93–100. 2020.
Article28. Kim KH, Lee JY, Wang KC. Secondary neurulation defects-1 : retained medullary cord. J Korean Neurosurg Soc. 63:314–320. 2020.
Article29. Kim KH, Wang KC, Lee JY. Enlargement of extraspinal cysts in spinal dysraphism : a reason for early untethering. J Korean Neurosurg Soc. 63:342–345. 2020.
Article30. Kirby ML, Lawson A, Stadt HA, Kumiski DH, Wallis KT, McCraney E, et al. Hensen’s node gives rise to the ventral midline of the foregut: implications for organizing head and heart development. Dev Biol. 253:175–188. 2003.
Article31. Lee JY, Kim KH, Park K, Wang KC. Retethering : a neurosurgical viewpoint. J Korean Neurosurg Soc. 63:346–357. 2020.
Article32. Lee JY, Kim SP, Kim SW, Park SH, Choi JW, Phi JH, et al. Pathoembryogenesis of terminal myelocystocele: terminal balloon in secondary neurulation of the chick embryo. Childs Nerv Syst. 29:1683–1688. 2013.
Article33. Lee JY, Pang D, Wang KC. Caudal Agenesis and Associated Spinal Cord Malformations. In : Di Rocco C, Pang D, Rutka JT, editors. Textbook of Pediatric Neurosurgery. Cham: Springer International Publishing;2020. p. 2557–2575.34. Lee JY, Park SH, Chong S, Phi JH, Kim SK, Cho BK, et al. Congenital dermal sinus and limited dorsal myeloschisis: “spectrum disorders” of incomplete dysjuction between cutaneous and neural ectoderms. Neurosurgery. 84:428–434. 2019.
Article35. Lee JY, Phi JH, Kim SK, Cho BK, Wang KC. Urgent surgery is needed when cyst enlarges in terminal myelocystoceles. Childs Nerv Syst. 27:2149–2153. 2011.
Article36. Li YC, Shin SH, Cho BK, Lee MS, Lee YJ, Hong SK, et al. Pathogenesis of lumbosacral lipoma: a test of the “premature dysjunction” theory. Pediatr Neurosurg. 34:124–130. 2001.
Article37. McLone DG, Naidich TP. Spinal dysraphism: experimental and clinical. In : Holtzman RN, Stein BM, editors. The tethered spinal cord. New York: Thieme;1985. p. 14–28.38. Mills CL, Bellairs R. Mitosis and cell death in the tail of the chick embryo. Anat Embryol (Berl). 180:301–308. 1989.
Article39. Moore KL, Persaud TVN, Torchia MG. The Developing Human-EBook: Clinically Oriented Embryology. Elsevier Health Sciences;2018. p. 193–223.40. Morota N, Ihara S, Ogiwara H. New classification of spinal lipomas based on embryonic stage. J Neurosurg Pediatr. 19:428–439. 2017.
Article41. Müller F, O’Rahilly R. The development of the human brain, the closure of the caudal neuropore, and the beginning of secondary neurulation at stage 12. Anat Embryol (Berl). 176:413–430. 1987.
Article42. Müller F, O’rahilly R. The primitive streak, the caudal eminence and related structures in staged human embryos. Cells Tissues Organs. 177:2–20. 2004.
Article43. Muecke EC. The role of the cloacal membrane in exstrophy: the first successful experimental study. J Urol. 92:659–667. 1964.44. Nour S, Kumar D, Dickson JA. Anorectal malformations with sacral bony abnormalities. Arch Dis Child. 64:1618–1620. 1989.
Article45. Ostling LR, Bierbrauer KS, Kuntz C 4th. Outcome, reoperation, and complications in 99 consecutive children operated for tight or fatty filum. World Neurosurg. 77:187–191. 2012.
Article46. Pang D. Sacral agenesis and caudal spinal cord malformations. Neurosurgery. 32:755–778. discussion 778-779. 1993.
Article47. Pang D, Chong S, Wang K. Secondary neurulation defects-1: thickened filum terminale, retained medullary cord. In : Di Rocco C, Pang D, Rutka JT, editors. Textbook of pediatric neurosurgery. ed 1. Switzerland: Springer;2020. p. 1–18.48. Pang D, Dias MS, Ahab-Barmada M. Split cord malformation: part I: a unified theory of embryogenesis for double spinal cord malformations. Neurosurgery. 31:451–480. 1992.49. Pang D, Zovickian J, Lee JY, Moes GS, Wang KC. Terminal myelocystocele: surgical observations and theory of embryogenesis. Neurosurgery. 70:1383–1404. discussion 1404-1405. 2012.50. Pang D, Zovickian J, Moes GS. Retained medullary cord in humans: late arrest of secondary neurulation. Neurosurgery. 68:1500–1519. discussion 1519. 2011.
Article51. Pang D, Zovickian J, Oviedo A. Long-term outcome of total and neartotal resection of spinal cord lipomas and radical reconstruction of the neural placode: part I-surgical technique. Neurosurgery. 65:511–528. discussion 528-529. 2009.52. Pang D, Zovickian J, Oviedo A, Moes GS. Limited dorsal myeloschisis: a distinctive clinicopathological entity. Neurosurgery. 67:1555–1579. discussion 1579-1580. 2010.
Article53. Sadler TW, Feldkamp ML. The embryology of body wall closure: relevance to gastroschisis and other ventral body wall defects. Am J Med Genet C Semin Med Genet. 148C:180–185. 2008.
Article54. Sala F, Barone G, Tramontano V, Gallo P, Ghimenton C. Retained medullary cord confirmed by intraoperative neurophysiological mapping. Childs Nerv Syst. 30:1287–1291. 2014.
Article55. Schoenwolf GC. Tail (end) bud contributions to the posterior region of the chick embryo. J Exp Zool. 201:227–245. 1977.
Article56. Schumacher S. Über Bildungs-und Rückbildungsvorgänge am Schwanzende des Medullarrohres bei älteren Hühnerembryonen mit besonderer Berücksichtigung des Auftretens eines „sekundären hinteren Neuroporus“. Z Mikrosk Anat Forsch. 13:269–327. 1928.57. Scott RM, Wolpert SM, Bartoshesky LE, Zimbler S, Karlin L. Segmental spinal dysgenesis. Neurosurgery. 22:739–744. 1988.
Article58. Shimokita E, Takahashi Y. Secondary neurulation: fate-mapping and gene manipulation of the neural tube in tail bud. Dev Growth Differ. 53:401–410. 2011.
Article59. Streit A, Lee KJ, Woo I, Roberts C, Jessell TM, Stern CD. Chordin regulates primitive streak development and the stability of induced neural cells, but is not sufficient for neural induction in the chick embryo. Development. 125:507–519. 1998.
Article60. Sur A, Sardar SK, Paria A. Caudal duplication syndrome. J Clin Neonatol. 2:101–102. 2013.
Article61. Uehara M, Yashiro K, Takaoka K, Yamamoto M, Hamada H. Removal of maternal retinoic acid by embryonic CYP26 is required for correct Nodal expression during early embryonic patterning. Genes Dev. 23:1689–1698. 2009.
Article62. Wang KC. Spinal dysraphism in the last two decades : what I have seen during the era of dynamic advancement. J Korean Neurosurg Soc. 63:272–278. 2020.
Article63. Wang KC, Lee JS, Kim K, Im YJ, Park K, Kim KH, et al. Do junctional neural tube defect and segmental spinal dysgenesis have the same pathoembryological background? Childs Nerv Syst. 36:241–250. 2020.
Article64. Wei Y, Mikawa T. Fate diversity of primitive streak cells during heart field formation in ovo. Dev Dyn. 219:505–513. 2000.
Article65. Wong ST, Kan A, Pang D. Limited dorsal spinal nondisjunctional disorders: Limited dorsal myeloschisis, congenital spinal dermal sinus tract, and mixed lesions. Di Rocco C, Pang D, Rutka JT. Textbook of Pediatric Neurosurgery. ed 1. Switzerland: Springer;2020. p. 1–64.66. Yamada S, Won DJ, Pezeshkpour G, Yamada BS, Yamada SM, Siddiqi J, et al. Pathophysiology of tethered cord syndrome and similar complex disorders. Neurosurg Focus. 23:E6. 2007.
Article67. Yamada S, Won DJ, Yamada SM. Pathophysiology of tethered cord syndrome: correlation with symptomatology. Neurosurg Focus. 16:E6. 2004.
Article68. Yamada S, Zinke DE, Sanders D. Pathophysiology of “tethered cord syndrome”. J Neurosurg. 54:494–503. 1981.
Article69. Yang J, Kim KH, Lee JY, Wang KC. Caudal duplication syndrome: a literature review and reappraisal of its pthoembryogenesis. Childs Nerv Syst. 2021; [Epub ahead of print].70. Zhang T, Zhang HL, Wang da J, Tang XB, Jia HM, Bai YZ, et al. Normal development of hindgut and anorectum in human embryo. Int J Colorectal Dis. 26:109–116. 2011.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Terminal Myelocystocele : Pathoembryogenesis and Clinical Features
- Perspectives : The Role of Clinicians in Understanding Secondary Neurulation
- Secondary Neurulation Defects-1 : Retained Medullary Cord
- Junctional Neurulation : A Junction between Primary and Secondary Neural Tubes
- Overview of Secondary Neurulation
