Mutant IDH1 Enhances Temozolomide Sensitivity via Regulation of the ATM/CHK2 Pathway in Glioma
- Affiliations
-
- 1Department of Neurosurgery, Second Affiliated Hospital of Harbin Medical University, Harbin, China
- KMID: 2514919
- DOI: http://doi.org/10.4143/crt.2020.506
Abstract
- Purpose
Isocitrate dehydrogenase 1 (IDH1) mutations are the most common genetic abnormalities in low-grade gliomas and secondary glioblastomas. Glioma patients with these mutations had better clinical outcomes. However, the effect of IDH1 mutation on drug sensitivity is still under debate.
Materials and Methods
IDH1-R132H mutant cells were established by lentivirus. IDH1-R132H protein expression was confirmed by western blot. The expression of ataxia telangiectasia mutated (ATM) signaling pathway and apoptosis-related proteins were detected by immunofluorescence and western blot. Temozolomide (TMZ) induced cell apoptosis was detected by flow cytometry. Tumor cell proliferation was detected by Cell Counting Kit-8. In vivo nude mice were used to confirm the in vitro roles of IDH1 mutation.
Results
We established glioma cell lines that expressed IDH1-R132H mutation stably. We found that TMZ inhibited glioma cells proliferation more significantly in IDH1 mutant cells compared to wild type. The IC50 of TMZ in IDH1-R132H mutant group was less than half that of wild-type group (p < 0.01). TMZ significantly induced more DNA damage (quantification of γH2AX expression in IDH1 mutation vs. wild type, p < 0.05) and apoptosis (quantification of AnnexinV+propidium iodide–cells in IDH1 mutation versus wild type, p < 0.01) in IDH1 mutant gliomas compared to wild-type gliomas. The ATM-associated DNA repair signal was impaired in IDH1 mutant cells. Inhibiting the ATM/checkpoint kinase 2DNA repair pathway further sensitized IDH1 mutant glioma cells to chemotherapy. We found that IDH1 mutation significantly inhibited tumor growth in vivo (the tumor size was analyzed statistically, p < 0.05). Moreover, we confirmed that gliomas with IDH1 mutation were more sensitive to TMZ in vivo compared to wild type significantly and the results were consistent with the in vitro experiment.
Conclusion
These results provide evidence that combination of TMZ and ATM inhibitor enhances the antitumor effect in IDH1 mutant gliomas.
Figure
-

Fig. 1 Isocitrate dehydrogenase 1 (IDH1) mutation inhibits temozolomide (TMZ) treated glioma cells proliferation. (A) IDH1 mutant cells that stably expressing IDH1-R132H mutants in Ln229 and U87 cell lines. IDH1-R132H expression was confirmed by western blot. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. (B, C) IDH1 mutant or IDH1 wild-type cells were treated with or without TMZ for 0 to 96 hours. Cell proliferation was detected by Cell Counting Kit-8. (D, E) TMZ dose-response curves of Ln229 and U87 cells with IDH1 mutation for 96 hours. (F) Statistical analysis of IC50 in Ln229 and U87 glioma cells. **p < 0.01.
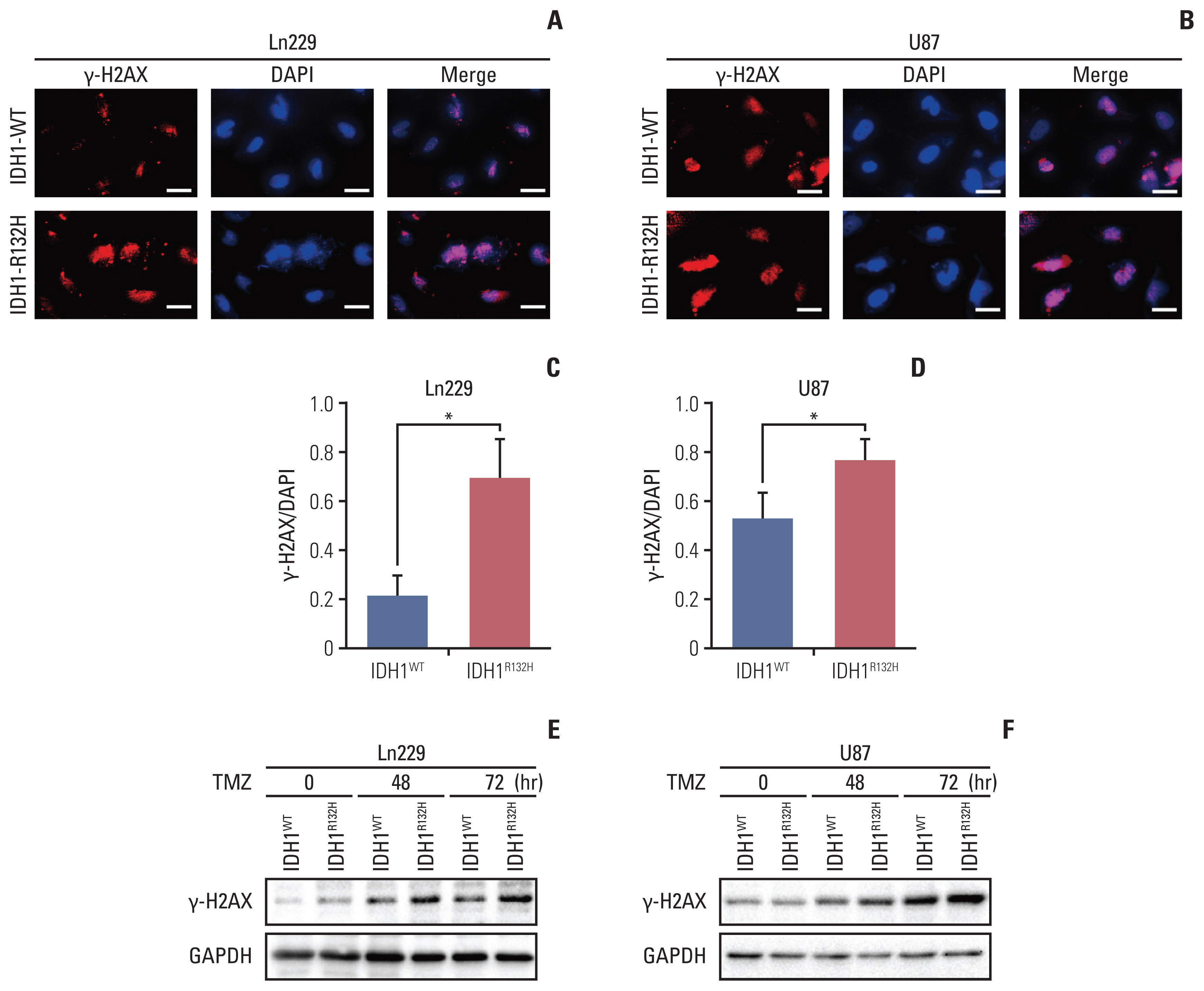
Fig. 2 Temozolomide (TMZ) induced more DNA damage in isocitrate dehydrogenase 1 (IDH1) mutation cells. (A, B) Immunofluorescence staining of γH2AX in glioma cells Ln229 and U87. Scale bars=100 μm. (C, D) Quantification of γH2AX is shown in A and B. *p < 0.05. (E, F) γH2AX expression in glioma cells Ln229 and U87 were detected by western blotting. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
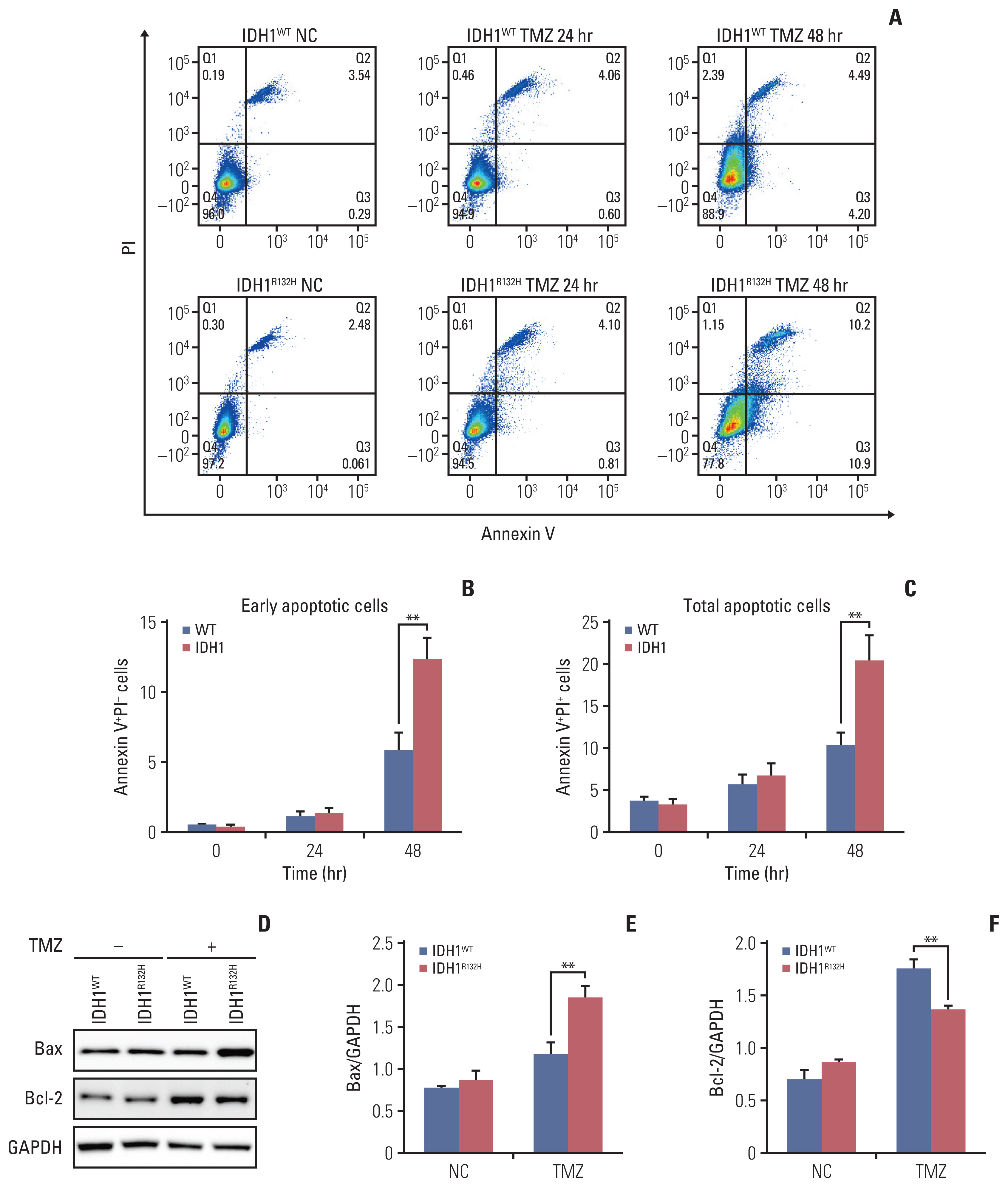
Fig. 3 Isocitrate dehydrogenase 1 (IDH1) mutant enhance temozolomide (TMZ) induced apoptosis. (A) Flow cytometry apoptosis analysis of IDH1 mutant and wild-type cells treated TMZ at different time points. (B) Statistical quantification of annexin V+propidium iodide (PI)− cells. (C) Statistical quantification of annexin V+PI+ cells. (D) Apoptosis-related protein Bax and Bcl-2 were detected by western blotting. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. (E, F) Quantification of relative expression of Bax and Bcl-2. **p < 0.01.

Fig. 4 Isocitrate dehydrogenase 1 (IDH1) mutation inhibits ataxia telangiectasia mutated (ATM)/checkpoint kinase 2 (CHK2) signaling triggered by temozolomide (TMZ). (A, B) The expression of ATM signaling including ATM, CHK2, p53, and their phosphorylated proteins in glioma cells treated with or without TMZ were detected by Western blotting. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
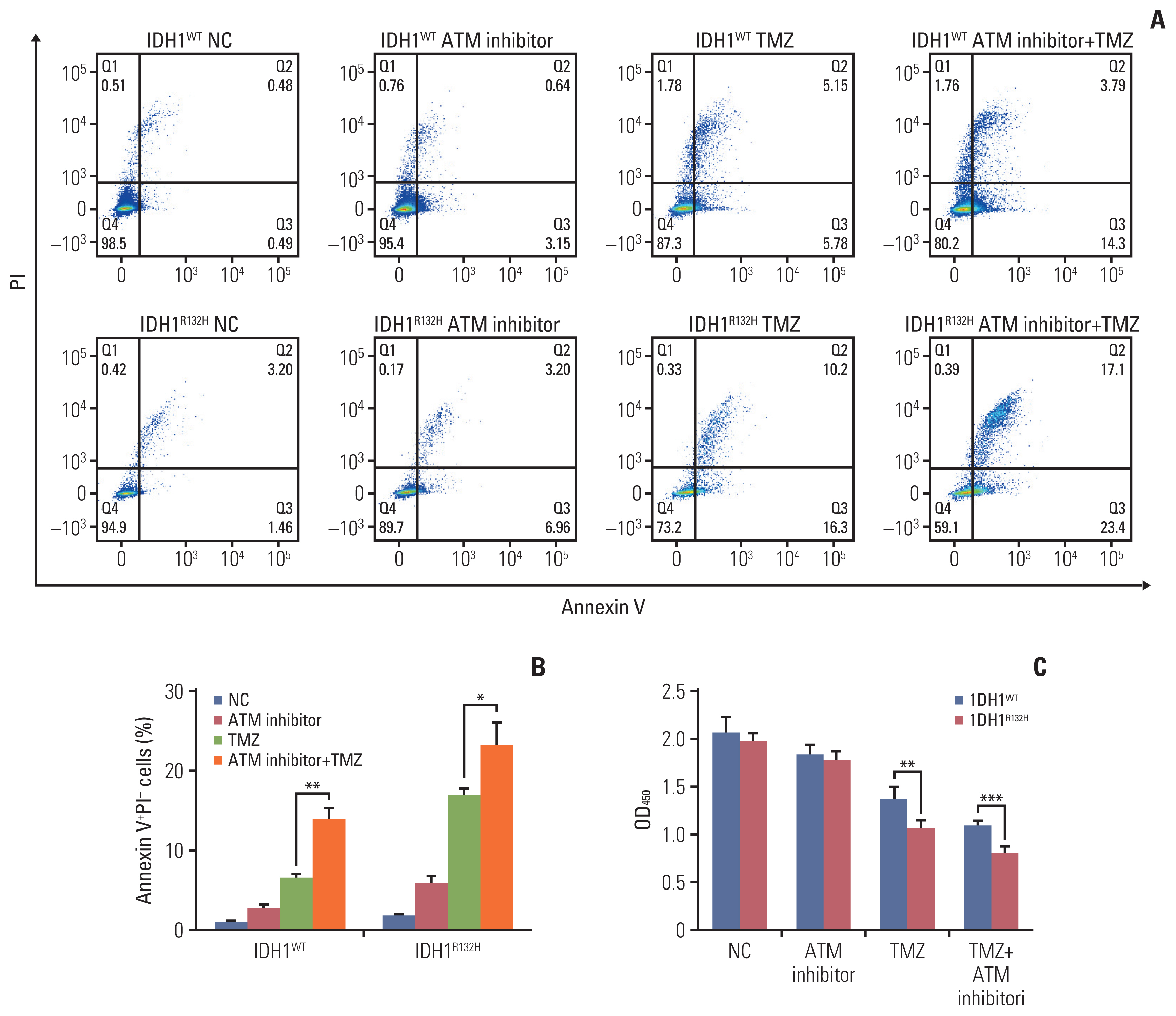
Fig. 5 Ataxia telangiectasia mutated (ATM) inhibitor enhanced the effect of temozolomide (TMZ) on tumor apoptosis. (A) Isocitrate dehydrogenase 1 (IDH1) mutation and wild-type gliomas were divided into four groups treated with negative control, ATM inhibitor, TMZ and ATM inhibitor+TMZ, after 72 hours, cell apoptosis was detected by flow cytometry. (B) Statistical quantification of annexin V+propidium iodide (PI)− cells. (C) IDH1 mutation and wild-type gliomas were treated with negative control, ATM inhibitor, TMZ and ATM inhibitor+TMZ, respectively. Cell proliferation was detected by Cell Counting Kit-8. *p < 0.05, **p < 0.01, ***p < 0.001.
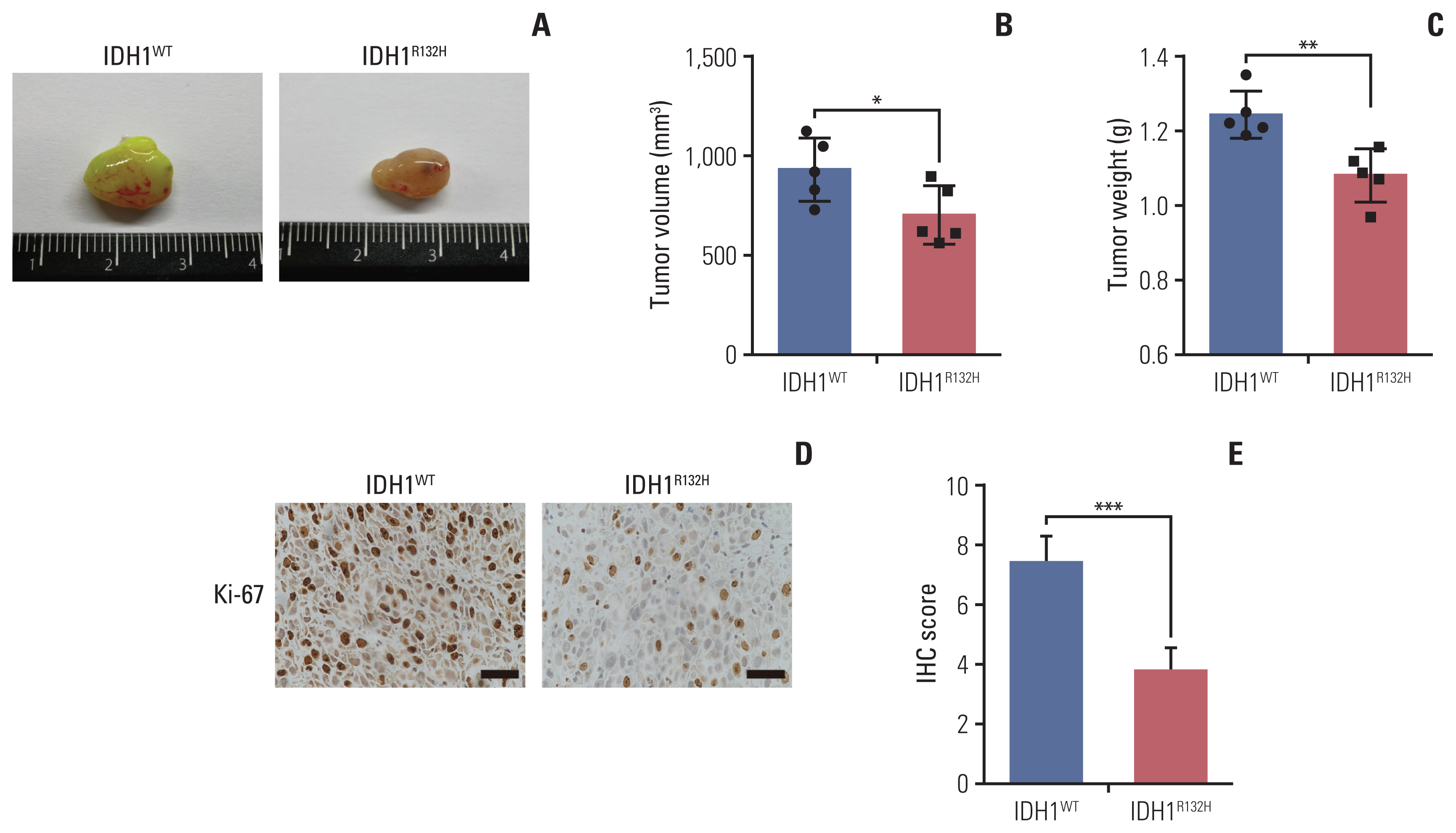
Fig. 6 The effect of isocitrate dehydrogenase 1 (IDH1) mutation on tumor in vivo. (A) The same number of IDH1 mutant and wild-type glioma cells were injected into the subcutaneous of 4–6 weeks nude mice, and the mice were killed and photographed in the fifth week. (B) The tumor size was detected and analyzed statistically. (C) The tumor weight was detected and analyzed statistically. (D) The tumor tissue was paraffin-embedded and stained with immunohistochemistry to detect the expression of Ki-67. Scale bars=25 μm. (E) The results of immunohistochemistry were scored and analyzed statistically. *p < 0.05, **p < 0.01, ***p < 0.001.
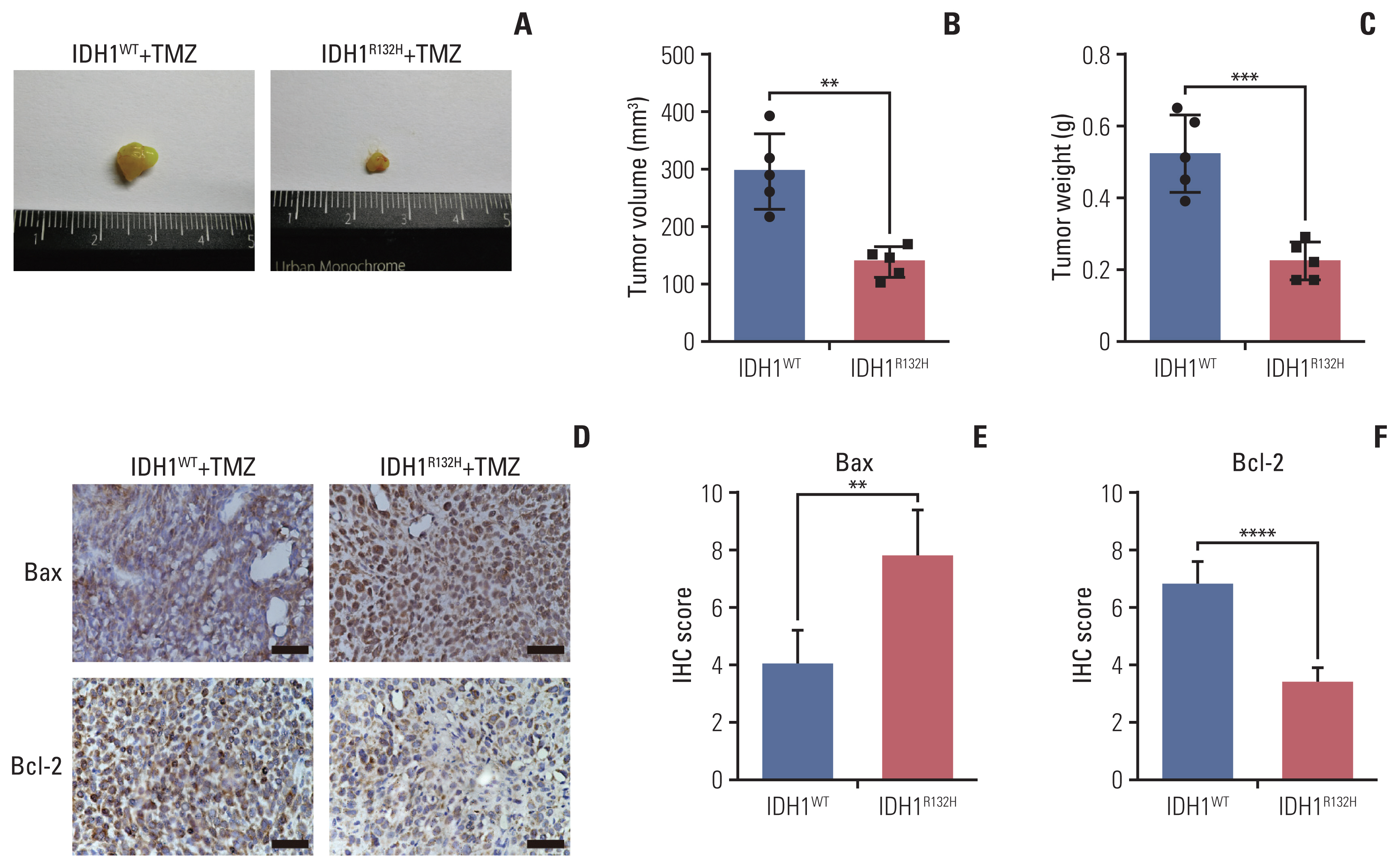
Fig. 7 The effect of temozolomide (TMZ) on isocitrate dehydrogenase 1 (IDH1) mutant glioma in vivo. (A) IDH1 mutant and wild-type glioma cells were subcutaneously injected into 4–6 weeks BALB/c nude mice. After transplanted for 4 weeks 20 mg/kg TMZ was administered intraperitoneally into mice for consecutive 7 days. All nude mice were killed in the sixth week. (B) The tumor volume was monitored and analyzed. (C) The tumor weight was monitored and analyzed. (D) Primary glioma sections from mouse models were stained with Bax and Bcl-2 for immunohistochemistry (IHC) assay. Scale bars=25 μm. (E, F) The results of IHC were scored and analyzed statistically. All results are expressed as the mean±standard deviation; n=5. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Reference
-
References
1. Louis DN, Perry A, Reifenberger G, von Deimling A, Figarella-Branger D, Cavenee WK, et al. The 2016 World Health Organization classification of tumors of the central nervous system: a summary. Acta Neuropathol. 2016; 131:803–20.
Article2. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005; 352:987–96.
Article3. Stupp R, Gander M, Leyvraz S, Newlands E. Current and future developments in the use of temozolomide for the treatment of brain tumours. Lancet Oncol. 2001; 2:552–60.
Article4. Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature. 2009; 462:739–44.
Article5. Sulkowski PL, Corso CD, Robinson ND, Scanlon SE, Purshouse KR, Bai H, et al. 2-Hydroxyglutarate produced by neomorphic IDH mutations suppresses homologous recombination and induces PARP inhibitor sensitivity. Sci Transl Med. 2017; 9:eaal2463.
Article6. Parsons DW, Jones S, Zhang X, Lin JC, Leary RJ, Angenendt P, et al. An integrated genomic analysis of human glioblastoma multiforme. Science. 2008; 321:1807–12.
Article7. Cancer Genome Atlas Research Network. Brat DJ, Verhaak RG, Aldape KD, Yung WK, Salama SR, et al. Comprehensive, integrative genomic analysis of diffuse lower-grade gliomas. N Engl J Med. 2015; 372:2481–98.8. SongTao Q, Lei Y, Si G, YanQing D, HuiXia H, XueLin Z, et al. IDH mutations predict longer survival and response to temozolomide in secondary glioblastoma. Cancer Sci. 2012; 103:269–73.
Article9. Ohba S, Mukherjee J, See WL, Pieper RO. Mutant IDH1-driven cellular transformation increases RAD51-mediated homologous recombination and temozolomide resistance. Cancer Res. 2014; 74:4836–44.
Article10. Matsuoka S, Rotman G, Ogawa A, Shiloh Y, Tamai K, Elledge SJ. Ataxia telangiectasia-mutated phosphorylates Chk2 in vivo and in vitro. Proc Natl Acad Sci U S A. 2000; 97:10389–94.
Article11. Helleday T, Petermann E, Lundin C, Hodgson B, Sharma RA. DNA repair pathways as targets for cancer therapy. Nat Rev Cancer. 2008; 8:193–204.
Article12. Lin L, Li Y, Liu M, Li Q, Liu Q, Li R. The Interleukin-33/ST2 axis promotes glioma mesenchymal transition, stemness and TMZ resistance via JNK activation. Aging (Albany NY). 2020; 12:1685–703.
Article13. Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009; 10:459–66.
Article14. Lin L, Cai J, Jiang C. Recent advances in targeted therapy for glioma. Curr Med Chem. 2017; 24:1365–81.
Article15. Yan H, Parsons DW, Jin G, McLendon R, Rasheed BA, Yuan W, et al. IDH1 and IDH2 mutations in gliomas. N Engl J Med. 2009; 360:765–73.16. Horbinski C. What do we know about IDH1/2 mutations so far, and how do we use it? Acta Neuropathol. 2013; 125:621–36.17. Cai J, Zhu P, Zhang C, Li Q, Wang Z, Li G, et al. Detection of ATRX and IDH1-R132H immunohistochemistry in the progression of 211 paired gliomas. Oncotarget. 2016; 7:16384–95.
Article18. Cai J, Yang P, Zhang C, Zhang W, Liu Y, Bao Z, et al. ATRX mRNA expression combined with IDH1/2 mutational status and Ki-67 expression refines the molecular classification of astrocytic tumors: evidence from the whole transcriptome sequencing of 169 samples samples. Oncotarget. 2014; 5:2551–61.19. Lu Y, Kwintkiewicz J, Liu Y, Tech K, Frady LN, Su YT, et al. Chemosensitivity of IDH1-mutated gliomas due to an impairment in PARP1-mediated DNA repair. Cancer Res. 2017; 77:1709–18.
Article20. Inoue S, Li WY, Tseng A, Beerman I, Elia AJ, Bendall SC, et al. Mutant IDH1 downregulates ATM and alters DNA repair and sensitivity to DNA damage independent of TET2. Cancer Cell. 2016; 30:337–48.
Article21. Cairncross JG, Wang M, Jenkins RB, Shaw EG, Giannini C, Brachman DG, et al. Benefit from procarbazine, lomustine, and vincristine in oligodendroglial tumors is associated with mutation of IDH. J Clin Oncol. 2014; 32:783–90.
Article22. Yang P, Zhang W, Wang Y, Peng X, Chen B, Qiu X, et al. IDH mutation and MGMT promoter methylation in glioblastoma: results of a prospective registry. Oncotarget. 2015; 6:40896–906.
Article23. Knizhnik AV, Roos WP, Nikolova T, Quiros S, Tomaszowski KH, Christmann M, et al. Survival and death strategies in glioma cells: autophagy, senescence and apoptosis triggered by a single type of temozolomide-induced DNA damage. PLoS One. 2013; 8:e55665.
Article24. Quiros S, Roos WP, Kaina B. Processing of O6-methylguanine into DNA double-strand breaks requires two rounds of replication whereas apoptosis is also induced in subsequent cell cycles. Cell Cycle. 2010; 9:168–78.
Article25. Burma S, Chen BP, Murphy M, Kurimasa A, Chen DJ. ATM phosphorylates histone H2AX in response to DNA double-strand breaks. J Biol Chem. 2001; 276:42462–7.
Article26. Han B, Cai J, Gao W, Meng X, Gao F, Wu P, et al. Loss of ATRX suppresses ATM dependent DNA damage repair by modulating H3K9me3 to enhance temozolomide sensitivity in glioma. Cancer Lett. 2018; 419:280–90.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Activation of ATM-dependent DNA Damage Signal Pathway by a Histone Deacetylase Inhibitor, Trichostatin A
- Identification of a New Selective Chemical Inhibitor of Mutant Isocitrate Dehydrogenase-1
- Temozolomide Drives Ferroptosis via a DMT1-Dependent Pathway in Glioblastoma Cells
- FoxD2-AS1 is a prognostic factor in glioma and promotes temozolomide resistance in a Oâ¶-methylguanine-DNA methyltransferase-dependent manner
- Analysis of Mutant Isocitrate Dehydrogenase 1 Immunoexpression, Ki-67 and Programmed Death Ligand 1 in Diffuse Astrocytic Tumours : Study of Single Center in Bandung, Indonesia
