T Cells Modified with CD70 as an Alternative Cellular Vaccine for Antitumor Immunity
- Affiliations
-
- 1Department of Microbiology and Immunology, College of Medicine, The Catholic University of Korea, Seoul, Korea
- 2Translational and Clinical Division, ViGenCell Inc., Seoul, Korea
- 3Catholic Hematopoietic Stem Cell Bank, College of Medicine, The Catholic University of Korea, Seoul, Korea
- KMID: 2504457
- DOI: http://doi.org/10.4143/crt.2019.721
Abstract
- Purpose
Successful tumor eradication primarily depends on generation and maintenance of a large population of tumor-reactive CD8 T cells. Dendritic cells (DCs) are well-known potent antigen-presenting cells and have applied to clinics as potent antitumor therapeutic agents. However, high cost and difficulty in obtaining sufficient amounts for clinical use are the crucial drawbacks of DC-based vaccines. Here, we aimed to develop T cell–based vaccine capable of eliciting potent antitumor therapeutic effects by providing effective costimulatory signals.
Materials and Methods
Antigenic peptide-loaded T cells transfected with retrovirus encoding costimulatory ligands CD70, CD80, OX40L, or 4-1BBL were assessed for antigen-specific CD8 T-cell responses and evaluated antitumor effects along with immunization of a mixture of synthetic peptides, poly-IC and anti-CD40 antibodies (TriVax).
Results
T cells expressing CD70 (CD70-T) exhibited similar level of stimulatory functionality and therapeutic efficacy as DCs. Moreover, CD70-T prime followed by TriVax booster heterologous vaccination elicited therapeutic antitumor effect against B16 melanoma where mediated by CD8 T cells but not CD4 T cells or natural killer cells. The combination with programmed death-ligand 1 blockade led to potent therapeutic efficacy which exhibited increased tumor-infiltrating CD8 T cells. CD70-T pulsed with multi-antigenic peptide generated multiple antigen-specific polyvalent CD8 T cells that were capable of inhibiting tumor growth effectively. Moreover, CD70-T vaccination resulted in higher expansion and migration of adoptively transferred T cells into tumor sites and elicits enhanced therapeutic effects with peptide-based booster immu-nization.
Conclusion
These results imply that T cells endowed with CD70 enable the design of effective vaccination strategies against solid cancer, which may overcome current limitations of DC-based vaccines.
Figure
-

Fig. 1. Genetically modified CD8 T-cell–based vaccination induces antigen-specific CD8 T-cell responses comparable to dendritic cell-based vaccination. Ex vivo activated splenocytes were transduced with recombinant retroviruses encoding mouse CD70, CD80, OX40L, and 4-1BBL (CD70-T, CD80-T, OX40L-T, and 4-1BBL-T, respectively). GFP was used as a mockcontrol (GFP-T). (A) The expression of transduced CD8 T cells was examined on day 3 post-transduction. (B) Cell apoptosis of the transduced were assessed by staining with annexin-V and 7-aminoactinomycin D (7-AAD) on day 3 post-transduction. (C) Proliferation of Pmel-1 T cells in response to genetically modified CD8 T cells. Pmel-1 T cells were labeled with 5 μM carboxyfluorescein succinimidyl ester and cultured with conditioned CD8 T cells as indicated, including DCs which were loaded with 1 μg/mL hgp10025 peptide. On day 3 post-co-culture, cell proliferation was measured by flow cytometry and analyzed using Modifit LT software (left panel). The results are the sum of the average proliferation index (PI) of Pmel-1 T cells from three independent experiments with standard deviation (SD, bars) of the means (right panel). PI: the sum of the cells in all generations divided by the computed number of parental cells present at the start of experiment. CFSE, carboxyfluorescein succinimidylester; ns, not significant; *p < 0.05. (D, F) B6 mice (2 per group) were immunized intravenously (i.v.) on day 0, 4, and 8 with 2×106 Trp1455/9M-loaded conditioned CD8 T cells as indicated. Antigen-loaded dendritic cells (DC) and GFP-T were used for comparison. (D) Seven days after the last immunization, frequency of antigen-specific CD8 T cells in spleen from individual mouse was evaluated by cell surface mobilization of CD107a/b and intracellular interferon γ (IFNγ) staining. The results are the sum of two independent experiments. Points and bars indicate values for each individual mouse and SD, respectively. (E) Freshly isolated CD8 T cells from pooled splenocytes in D were evaluated for antigen-induced IFNγ secretions by EliSpot against un-pulsed (EL4) and peptide-pulsed EL4 (EL4/Trp1455), and B16 tumor cells. Results represent the average number of spots from duplicate wells with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, ***p < 0.001). (F) Therapeutic efficacy of Trp1455/9M-loaded conditioned CD8 T cells against 3-day-established B16 melanoma. B6 mice (5 per group) were inoculated subcutaneously (s.c.) with 1×105 B16 cells on day 0 and received 2×106 Trp1455/9M-loaded DCs or CD8 T cells on day 3, 7, and 11 (vertical arrows). Non-vaccinated mice (No vax) were included as controls. Tumor size was determined in individual mice by measurements of two opposing diameters and are presented as tumor areas in mm2. Points and bars indicate mean for each group of mice and SD, respectively. p-values were calculated using 2-way ANOVA test (ns, not significant; *p < 0.05, **p < 0.01). These experiments were repeated twice with similar results.

Fig. 2. Peptide-based booster immunization after priming with CD70-T cells drives enhanced CD8 T-cell responses. B6 mice (2 per group) were immunized intravenously on day 0 with 2×106 Trp1455/9M-loaded CD70-T cells; 7 days later, the mice received a booster immunization with various combination of 100 μg of Trp1455/9M peptide, 50 μg of poly-IC, and 50 μg anti-CD40, anti–4-1BB, or anti-OX40 antibodies. (A) Seven days after last immunization, the frequency of antigen-specific CD8 T cells in spleen was evaluated as in Fig. 1D. A representative dot plot analysis for one mouse of each group is presented (left panel). Numbers in each rectangular gate represent the percentage of interferon γ (IFNγ) and cell surface CD107a/b double-positive cells of all CD8 T cells. The results are the sum of two independent experiments. Points and bars indicate values for each individual mouse and standard deviation (SD), respectively. (B) Antigen-induced IFNγ-secretions with freshly isolated CD8 T cells from pooled splenocytes in A were evaluated as in Fig. 1E. Results represent the average number of spots from duplicate wells with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, **p < 0.01, ***p < 0.001). These experiments were repeated twice with similar results.
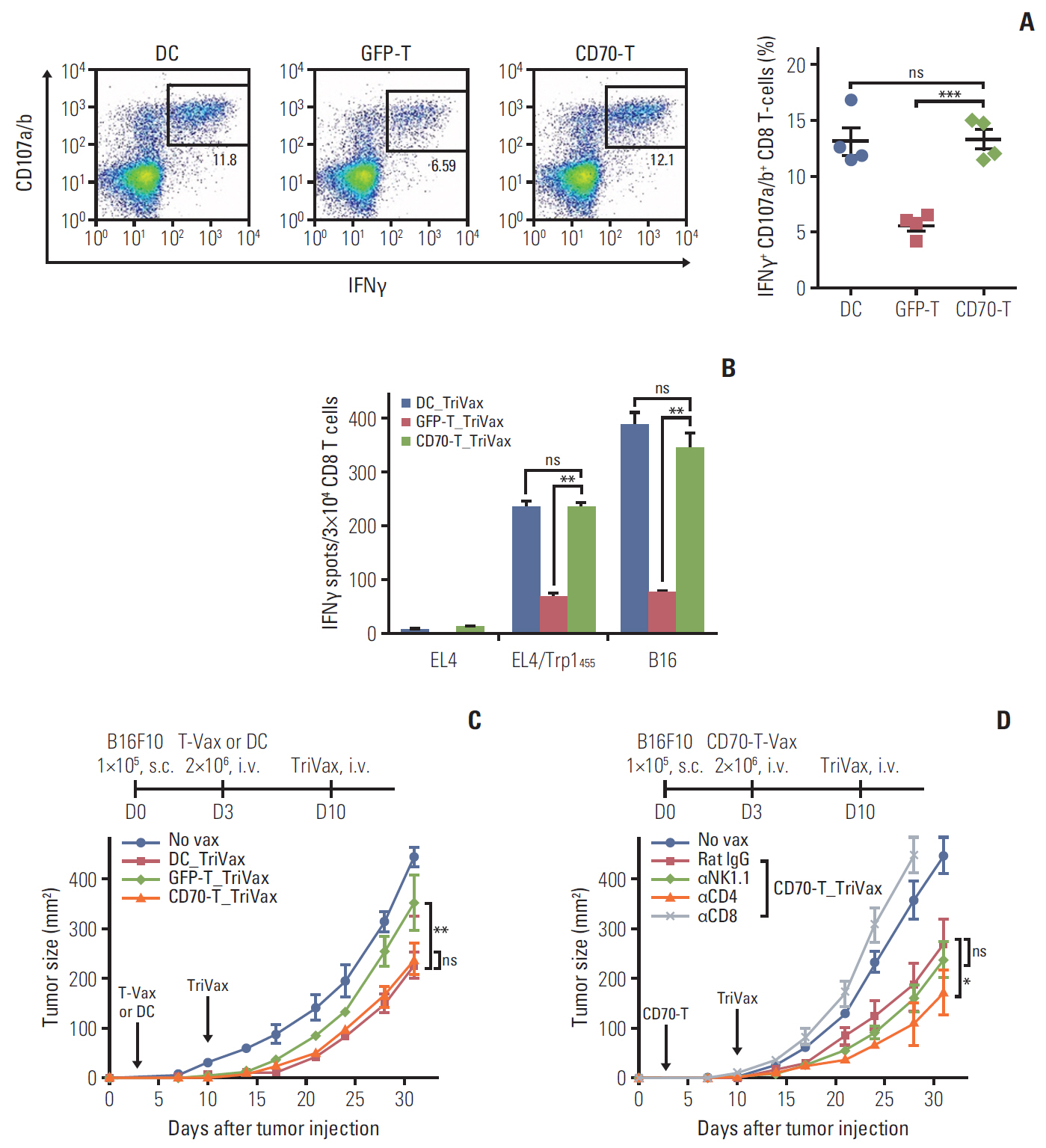
Fig. 3. Therapeutic antitumor effects of CD70-T prime_TriVax boost vaccination against established B16 melanoma. B6 mice (2 per group) were immunized intravenously (i.v.) on day 0 with 2×106 Trp1455/9M-loaded GFP-T or CD70-T cells; 7 days later, the mice received a booster immunization with combination of 100 μg of Trp1455/9M peptide, 50 μg of poly-IC, and 50 μg anti-CD40 antibodies (GFP-T_TriVax and CD70-T_TriVax, respectively). Trp1455/9M-loaded dendritic cells prime_TriVax boost (DC_TriVax) were included for comparison. (A) Seven days after last immunization, the frequency of antigen-specific CD8 T cells in spleen was evaluated as in Fig. 1D. A representative dot plot analysis for one mouse of each group is presented (left panel). Numbers in each rectangular gate represent the percentage of interferon γ (IFNγ) and cell surface CD107a/b double-positive cells of all CD8 T cells. The results are the sum of two independent experiments. Points and bars indicate values for each individual mouse and standard deviation (SD), respectively. (B) Antigen-induced IFNγ-secretions with freshly isolated CD8 T cells from pooled splenocytes in A were evaluated as in Fig. 1E. Results represent the average number of spots from duplicate wells with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; **p < 0.01, ***p < 0.001). (C) Therapeutic efficacy of CD70-T_TriVax vaccination against 3-day-established B16 melanoma. B6 mice (5 per group) were inoculated subcutaneously (s.c.) with 1×105 B16 cells on day 0 and received 2×106 Trp1455/9M-loaded GFP-T or CD70-T cells on day 3. After 7 days, the mice received a TriVax booster immunization (vertical arrows). Non-vaccinated mice (No vax) and DC_TriVax were included for comparison. (D) Effector mechanism of CD70-T_TriVax vaccination involved in antitumor effects. B6 mice (5 per group) were administered intraperitoneally with indicated antibodies –1 and –3 days before each immunization. No vax and Rat IgG-treated mice were included as controls. Tumor sizes were determined in individual mice by measurements of two opposing diameters and are presented as tumor areas in mm2. Points and bars indicate the mean for each group of mice and SD, respectively. p-values were calculated using 2-way ANOVA test (ns, not significant; *p < 0.05, **p < 0.01). These experiments were repeated twice with similar results.
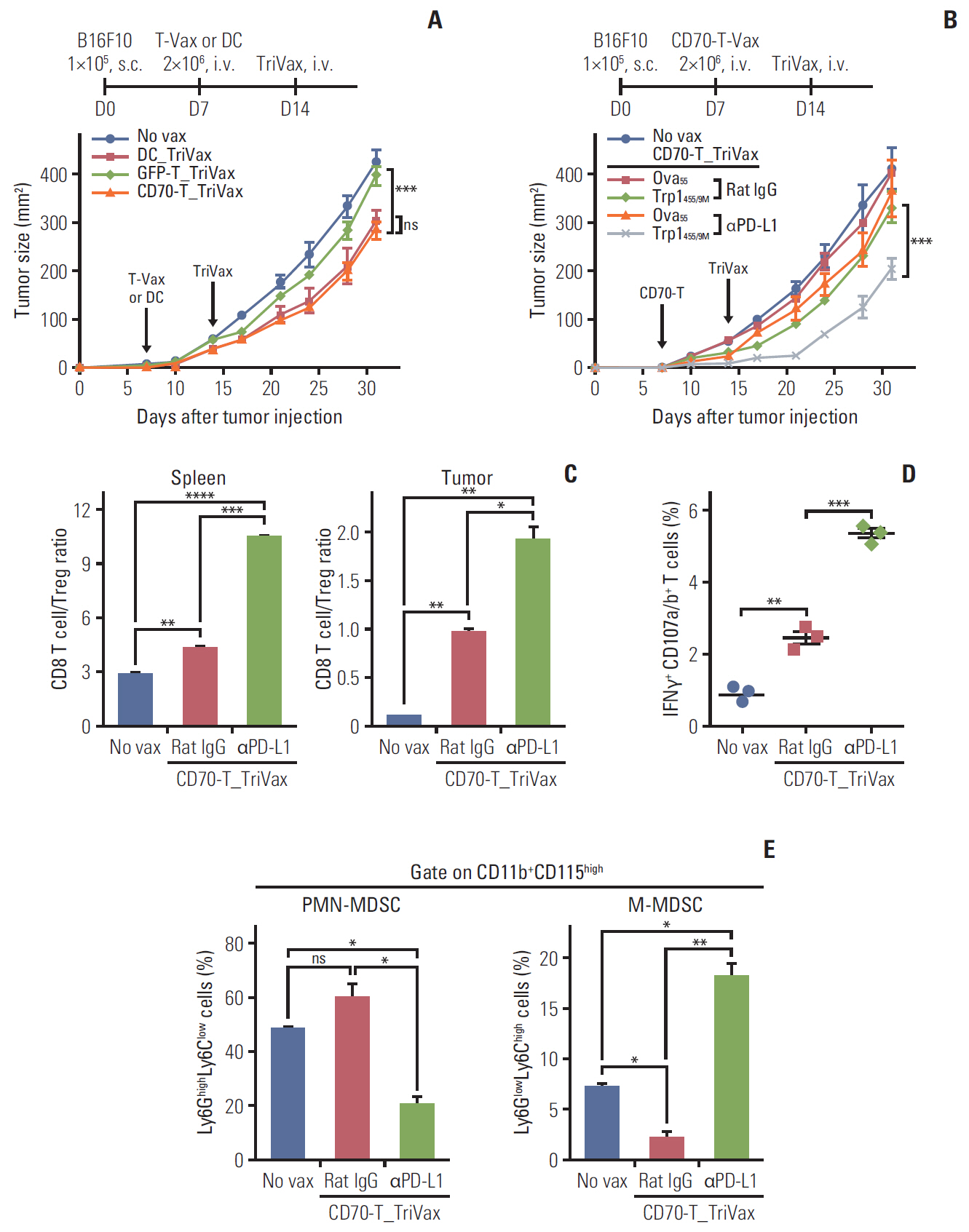
Fig. 4. Programmed death-ligand 1 (PD-L1) blockade augments the therapeutic efficacy of CD70-T_TriVax vaccination. (A) Therapeutic efficacy of CD70-T_TriVax vaccination against advanced B16 melanoma. B6 mice (7 per group) were inoculated subcutaneously on day 0 with 1×105 B16 cells, followed by vaccination with 2×106 GFP-T or CD70-T cells on day 7, and TriVax on day 14 (vertical arrows). Non-vaccinated mice (No vax) and DC_TriVax were included for comparison. (B) PD-L1 blockade enhanced the efficacy of CD70-T_TriVax vaccination regimen. B6 mice (5 per group) were inoculated subcutaneously (s.c.) on day 0 with 1×105 B16 cells and were immunized as described in A. Rat IgG (as controls) and anti–PD-L1 antibodies were administered intraperitoneally on days +1 and +3 after each immunization. No vaccinated (No vax) and Ova55-loaded CD70-T_TriVax vaccinated mice (Ova55) were included for comparison. Tumor sizes were determined in individual mice by measurements of two opposing diameters and are presented as tumor areas in mm2. Points and bars indicate the mean for each group of mice and standard deviation (SD), respectively. p-values were calculated using 2-way ANOVA test (ns, not significant; ***p < 0.001). (C-E) In a parallel with B, mice (3 per group) were sacrificed on day 21, and the CD8 T-cell responses and immune cell populations were evaluated. Cells from disaggregated tissues of spleen and tumor were analyzed for the composition of various subsets of immune cells. (C) CD8 T cell/Treg ratio as measured by percentage of CD8+ T cells per percentage of CD4+ Foxp3+ T cells cells in each group. (D) The frequency of antigen-specific CD8 T cells in spleen was evaluated as in Fig. 1D. Points and SD indicate values for each individual mouse and SD, respectively. ns, not significant; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. (E) Quantification of Ly6GlowLy6Chigh monocytic myeloid-derived suppressor cells (M-MDSC) and Ly6GhighLy6Clow granulocytic polymorphonuclear MDSC (PMN-MDSC) gated on CD11b+CD115highmyeloid cells in tumor sites. Results represent the average percentage of the gated cells from individual mice with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, **p < 0.01. These experiments were repeated twice with similar results.
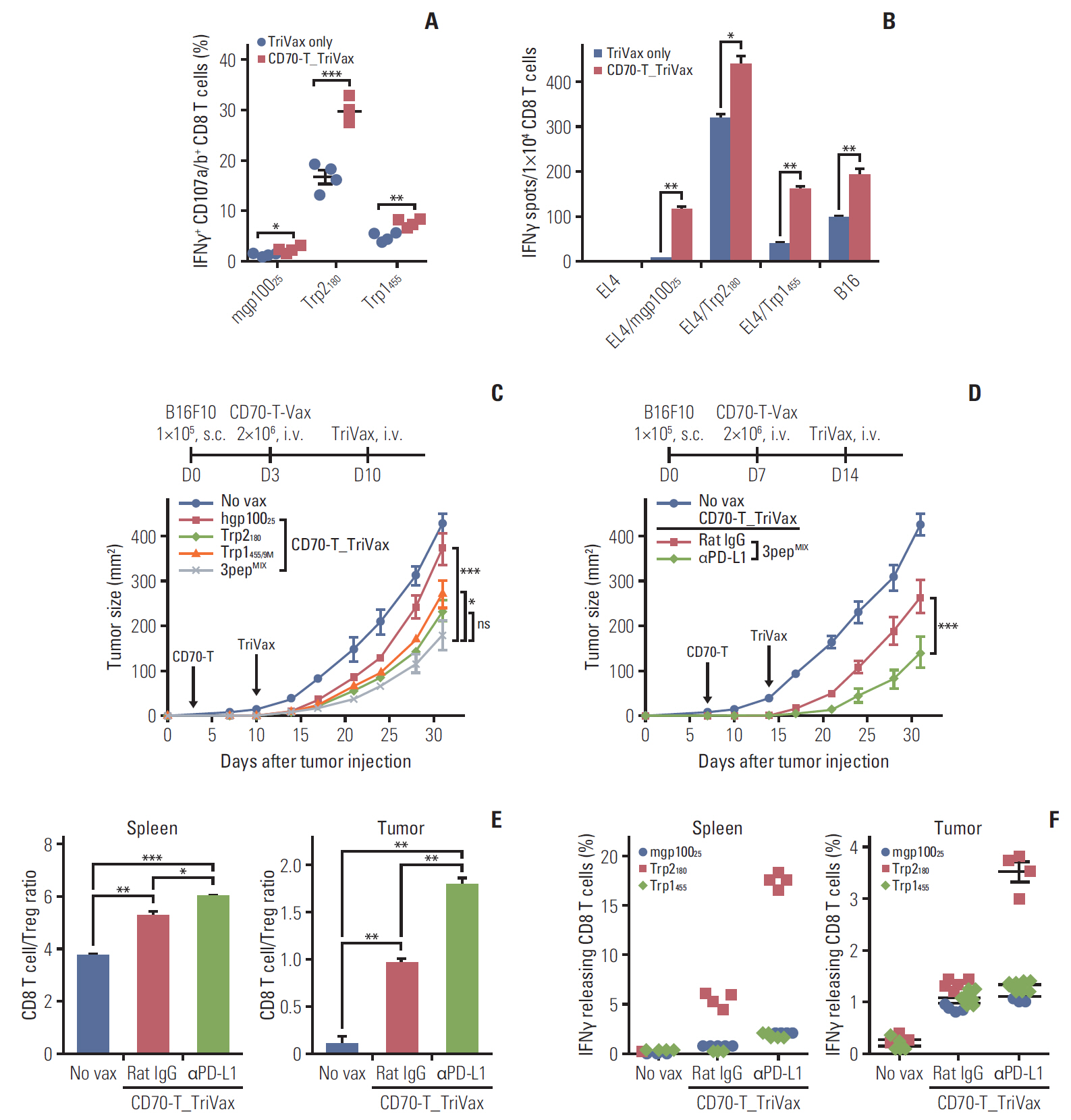
Fig. 5. Multi-epitope-loaded CD70-T cells priming followed by TriVax booster immunization induces concurrent effective antitumor CD8 T-cell responses. B6 mice (2 per group) were immunized intravenously (i.v.) with CD70-T cells that were loaded with mixture of Trp1455/9M, Trp2180, and hgp10025 peptides; 7 days later, the mice received 3pepMIXTriVax. (A) The frequency of antigen-specific CD8 T cells in spleen was evaluated as in Fig. 1D. Points and standard deviation (SD) indicate values for each individual mouse and SD, respectively. (B) Antigen-induced interferon γ (IFNγ)-secretions with freshly isolated CD8 T cells from pooled splenocytes in A were evaluated as in Fig. 1E. Results represent the average number of spots from duplicate wells with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, **p < 0.01, ***p < 0.001). (C) Therapeutic effectiveness of multi-epitope–loaded CD70-T_TriVax immunization against 3-day–established B16 melanoma. B6 mice (5 per group) were inoculated subcutaneously on day 0 with 1×105 B16 cells and were immunized i.v. on day 3 and 10 (vertical arrow) in combination with CD70-T or TriVax using either individual peptide or mixture of Trp1455/9M, Trp2180, and hgp10025 (3pepMIX) peptides as indicated. (D) Programmed death-1 blockade enhance the therapeutic efficacy of 3pepMIXCD70-T_TriVax immunization against advanced B16 tumors. B6 mice (7 per group) were inoculated subcutaneously (s.c.) on day 0 with 1×105 B16 cells and were immunized as described in Fig. 4B. Rat IgG (as controls) and anti–PD-L1 antibodies were administered intraperitoneally on days +1 and +3 after each immunization. Non-vaccinated mice (No vax) was included as a control. Tumor sizes were determined in individual mice by measurements of two opposing diameters and are presented as tumor areas on mm2. Points and bars indicate the mean for each group of mice and SD, respectively. p-values were calculated using 2-way ANOVA test (ns, not significant; *p < 0.05, ***p < 0.001). (E, F) In a parallel with D, mice (4 per group) were sacrificed on day 21, and the CD8 T-cell responses and immune cell populations were evaluated as described in Fig. 4C and D. (E) CD8 T-cell/Treg ratio as measured by percentage of CD8+ T cells per percentage of CD4+ Foxp3+ T cells cells in each group. Results represent the ave-rage percentage of the gated cells from individual mice with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (*p < 0.05, **p < 0.01, ***p < 0.001). (F) The frequency of antigen-specific CD8 T cells of spleen and tumor site was evaluated by intracellular IFNγ staining. Points and bars indicate values for each individual mouse and SD, respectively. These experiments were repeated twice with similar results.

Fig. 6. Effects of CD70-T vaccination in antitumor efficacy of adoptive T-cell therapy. (A-C) B6 mice (2 per group) were adoptively transferred 5×106 naïve Pmel-1 T cells on day 0, followed by immunization on day 1 with 3×106 hgp10025-pulsed GFP-T and CD70-T cells. Pmel-1 adoptive cell transfer (ACT) alone (ACT only) and dendritic cell vaccinated mice (DC) were included for comparison. (A) Seven days after the immunization, in vivo expansion of Pmel-1 T cells (CD90.1+CD8 T cells) in spleen was assessed. (B) The frequency of mgp10025-specific CD8 T cells in A was evaluated as in Fig. 1D. The results are the sum of two independent experiments with two per group. Points and bars indicate values for each individual mouse and SD, respectively. (C) Antigen-induced interferon γ (IFNγ)-secretions with freshly isolated CD8 T cells from pooled splenocytes in A were evaluated as in Fig. 1E. Results represent the average number of spots from duplicate wells with SD (bars) of the means. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, **p < 0.01, ***p < 0.001). (D, F) Antitumor effects of CD70-T immunization after Pmel-1 ACT. B6 mice (5 per group) were inoculated subcutaneously (s.c.) on day 0 with 1×105 B16 cells and received 5×106 Pmel-1 cells on day 3 followed by vaccination with hgp10025-pulsed GFP-T and CD70-T cells once (on day 4; D) or thrice (on day 4, 7, and 10; F, vertical arrows). For CD70-T_TriVax regimen, TriVax was administered intravenously (i.v.) 5 days after CD70-T vaccination. Non-vaccinated mice (No vax), Pmel-1 ACT alone (ACT only) and DC vaccinated mice (DC) were included for comparison. Tumor sizes were determined in individual mice by measurements of two opposing diameters and are presented as tumor areas in mm2. Points and bars indicate the mean for each group of mice and SD, respectively. p-values were calculated using 2-way ANOVA test (ns, not significant; ***p < 0.001). (E, G) In a parallel with D and F, respectively, mice (4 per group) were sacrificed on day 21, and the frequency of Pmel-1 T cells in spleen and tumor site was evaluated. Points and SD indicate values for each individual mouse and SD, respectively. p-values were calculated using unpaired Student’s t test (ns, not significant; *p < 0.05, **p < 0.01, ***p < 0.001). These experiments were repeated twice with similar results.
Reference
-
References
1. Croft M. Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol. 2003; 3:609–20.
Article2. Thery C, Amigorena S. The cell biology of antigen presentation in dendritic cells. Curr Opin Immunol. 2001; 13:45–51.3. Cho HI, Jung SH, Sohn HJ, Celis E, Kim TG. An optimized peptide vaccine strategy capable of inducing multivalent CD8(+) T cell responses with potent antitumor effects. Oncoimmunology. 2015; 4:e1043504.4. Liu Y, Zhang X, Zhang W, Chen Z, Chan T, Ali K, et al. Adenovirus-mediated CD40 ligand gene-engineered dendritic cells elicit enhanced CD8(+) cytotoxic T-cell activation and antitumor immunity. Cancer Gene Ther. 2002; 9:202–8.
Article5. Mende I, Engleman EG. Breaking tolerance to tumors with dendritic cell-based immunotherapy. Ann N Y Acad Sci. 2005; 1058:96–104.
Article6. Steinman RM, Witmer-Pack M, Inaba K. Dendritic cells: antigen presentation, accessory function and clinical relevance. Adv Exp Med Biol. 1993; 329:1–9.
Article7. Turnis ME, Rooney CM. Enhancement of dendritic cells as vaccines for cancer. Immunotherapy. 2010; 2:847–62.
Article8. Shin CA, Cho HW, Shin AR, Sohn HJ, Cho HI, Kim TG. Co-expression of CD40L with CD70 or OX40L increases B-cell viability and antitumor efficacy. Oncotarget. 2016; 7:46173–86.
Article9. Guo S, Xu J, Denning W, Hel Z. Induction of protective cytotoxic T-cell responses by a B-cell-based cellular vaccine requires stable expression of antigen. Gene Ther. 2009; 16:1300–13.
Article10. Park MY, Kim HS, Woo SJ, Kim CH, Park JS, Sohn HJ, et al. Efficient antitumor immunity in a murine colorectal cancer model induced by CEA RNA-electroporated B cells. Eur J Immunol. 2008; 38:2106–17.
Article11. Kim YJ, Ko HJ, Kim YS, Kim DH, Kang S, Kim JM, et al. alpha-Galactosylceramide-loaded, antigen-expressing B cells prime a wide spectrum of antitumor immunity. Int J Cancer. 2008; 122:2774–83.12. Chung Y, Kim BS, Kim YJ, Ko HJ, Ko SY, Kim DH, et al. CD1d-restricted T cells license B cells to generate long-lasting cytotoxic antitumor immunity in vivo. Cancer Res. 2006; 66:6843–50.
Article13. Himoudi N, Morgenstern DA, Yan M, Vernay B, Saraiva L, Wu Y, et al. Human gammadelta T lymphocytes are licensed for professional antigen presentation by interaction with opsonized target cells. J Immunol. 2012; 188:1708–16.14. Kennedy R, Undale AH, Kieper WC, Block MS, Pease LR, Celis E. Direct cross-priming by th lymphocytes generates memory cytotoxic T cell responses. J Immunol. 2005; 174:3967–77.
Article15. Adamopoulou E, Diekmann J, Tolosa E, Kuntz G, Einsele H, Rammensee HG, et al. Human CD4+ T cells displaying viral epitopes elicit a functional virus-specific memory CD8+ T cell response. J Immunol. 2007; 178:5465–72.
Article16. Park HM, Sohn HJ, Kim YJ, Cho HI, Kim TG. CD4 T-cells transduced with CD80 and 4-1BBL mRNA induce long-term CD8 T-cell responses resulting in potent antitumor effects. Vaccine. 2014; 32:6919–26.
Article17. Croft M. Costimulation of T cells by OX40, 4-1BB, and CD27. Cytokine Growth Factor Rev. 2003; 14:265–73.
Article18. Driessens G, Kline J, Gajewski TF. Costimulatory and coinhibitory receptors in anti-tumor immunity. Immunol Rev. 2009; 229:126–44.
Article19. Johnson BD, Gershan JA, Natalia N, Zujewski H, Weber JJ, Yan X, et al. Neuroblastoma cells transiently transfected to simultaneously express the co-stimulatory molecules CD54, CD80, CD86, and CD137L generate antitumor immunity in mice. J Immunother. 2005; 28:449–60.
Article20. Overwijk WW, Theoret MR, Finkelstein SE, Surman DR, de Jong LA, Vyth-Dreese FA, et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state of self-reactive CD8+ T cells. J Exp Med. 2003; 198:569–80.
Article21. Garrigan K, Moroni-Rawson P, McMurray C, Hermans I, Abernethy N, Watson J, et al. Functional comparison of spleen dendritic cells and dendritic cells cultured in vitro from bone marrow precursors. Blood. 1996; 88:3508–12.
Article22. Datta J, Terhune JH, Lowenfeld L, Cintolo JA, Xu S, Roses RE, et al. Optimizing dendritic cell-based approaches for cancer immunotherapy. Yale J Biol Med. 2014; 87:491–518.23. Zarnani AH, Torabi-Rahvar M, Bozorgmehr M, Zareie M, Mojtabavi N. Improved efficacy of a dendritic cell-based vaccine against a murine model of colon cancer: the helper protein effect. Cancer Res Treat. 2015; 47:518–26.
Article24. Cho HI, Kim EK, Park SY, Lee SK, Hong YK, Kim TG. Enhanced induction of anti-tumor immunity in human and mouse by dendritic cells pulsed with recombinant TAT fused human survivin protein. Cancer Lett. 2007; 258:189–98.
Article25. Watts TH, Bertram EM, Bukczynski J, Wen T. T cell costimulatory molecules in anti-viral immunity: Potential role in immunotherapeutic vaccines. Can J Infect Dis. 2003; 14:221–9.
Article26. Arens R, Schepers K, Nolte MA, van Oosterwijk MF, van Lier RA, Schumacher TN, et al. Tumor rejection induced by CD70-mediated quantitative and qualitative effects on effector CD8+ T cell formation. J Exp Med. 2004; 199:1595–605.
Article27. Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol. 2007; 8:239–45.
Article28. Duraiswamy J, Kaluza KM, Freeman GJ, Coukos G. Dual blockade of PD-1 and CTLA-4 combined with tumor vaccine effectively restores T-cell rejection function in tumors. Cancer Res. 2013; 73:3591–603.
Article29. Mumprecht S, Schurch C, Schwaller J, Solenthaler M, Ochsenbein AF. Programmed death 1 signaling on chronic myeloid leukemia-specific T cells results in T-cell exhaustion and disease progression. Blood. 2009; 114:1528–36.
Article30. Pilon-Thomas S, Mackay A, Vohra N, Mule JJ. Blockade of programmed death ligand 1 enhances the therapeutic efficacy of combination immunotherapy against melanoma. J Immunol. 2010; 184:3442–9.
Article31. Landsberg J, Kohlmeyer J, Renn M, Bald T, Rogava M, Cron M, et al. Melanomas resist T-cell therapy through inflammation-induced reversible dedifferentiation. Nature. 2012; 490:412–6.
Article32. Mehrotra S, Britten CD, Chin S, Garrett-Mayer E, Cloud CA, Li M, et al. Vaccination with poly(IC:LC) and peptide-pulsed autologous dendritic cells in patients with pancreatic cancer. J Hematol Oncol. 2017; 10:82.
Article33. Aravantinou M, Frank I, Hallor M, Singer R, Tharinger H, Kenney J, et al. PolyICLC exerts pro- and anti-HIV effects on the DC-T cell milieu in vitro and in vivo. PLoS One. 2016; 11:e0161730.
Article34. Johnson P, Challis R, Chowdhury F, Gao Y, Harvey M, Geldart T, et al. Clinical and biological effects of an agonist anti-CD40 antibody: a Cancer Research UK phase I study. Clin Cancer Res. 2015; 21:1321–8.
Article35. Bajor DL, Mick R, Riese MJ, Huang AC, Sullivan B, Richman LP, et al. Long-term outcomes of a phase I study of agonist CD40 antibody and CTLA-4 blockade in patients with metastatic melanoma. Oncoimmunology. 2018; 7:e1468956.
Article36. Kumai T, Fan A, Harabuchi Y, Celis E. Cancer immunotherapy: moving forward with peptide T cell vaccines. Curr Opin Immunol. 2017; 47:57–63.
Article37. Hirayama M, Nishimura Y. The present status and future prospects of peptide-based cancer vaccines. Int Immunol. 2016; 28:319–28.
Article38. Peng M, Mo Y, Wang Y, Wu P, Zhang Y, Xiong F, et al. Neoantigen vaccine: an emerging tumor immunotherapy. Mol Cancer. 2019; 18:128.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- CD70 Expression on Spodoptera Frugiperda(Sf9) Cells by Baculovirus to Induce CD27 Stimulation in Mouse B Cells
- Expression of CD70 during Thymus Regeneration in the Rat
- Influence of Immunity Induced at Priming Step on Mucosal Immunization of Heterologous Prime-Boost Regimens
- Analysis of Antitumor Mechanism of Intravesical BCG Therapy in Tumorigenesis(II) Study about Memory T Cells and Lymphocyte Homing Receptor Positive Cells
- Inflammasomes in antiviral immunity: clues for influenza vaccine development
