Augmented Serum Amyloid A1/2 Mediated by TNF-induced NF-κB in Human Serous Ovarian Epithelial Tumors
- Affiliations
-
- 1Department of Biochemistry and Cancer Biology, Meharry Medical College, Nashville, TN 37208, USA. dson@mmc.edu
- 2Department of Pharmaceutical Sciences, College of Pharmacy, Florida A&M University, Tallahassee, FL 32301, USA.
- 3Department of Obstetrics and Gynecology, Vanderbilt-Ingram Cancer Center, Vanderbilt University Medical Center, Nashville, TN 37232, USA.
- KMID: 2376870
- DOI: http://doi.org/10.4110/in.2017.17.2.121
Abstract
- Tumor necrosis factor-α (TNF) is well known to be involved in the immune system and ovarian inflammation. Ovarian cancer is an inflammation-related malignancy that lacks early screening strategies, resulting in late diagnosis followed by high mortality. Based on our previous data, TNF induced abundant serum amyloid A (SAA), an acute phase protein linked to inflammation, in ovarian granulosal cells. To date, the regulation and expression of SAA in ovarian cancer is not fully elucidated. Here, we investigated the relationship between TNF and SAA by comparing human normal ovarian tissues and serous ovarian tumors. We found that SAA1/2 was significantly expressed in tumor tissues, but no or trace expression levels in normal tissues. TNF was also significantly upregulated in ovarian tumor tissues compared to normal tissues. Moreover, TNF significantly increased SAA1/2 levels in human ovarian cancer cell lines, OVCAR-3 and SKOV-3, in a time-dependent manner. Since the SAA1 promoter contains two nuclear factor (NF)-κB sites, we examined whether TNF regulates SAA1 promoter activity. Deletion analysis revealed that the proximal NF-κB site (−95/−85) played a critical role in regulating TNF-induced SAA1 promoter activity. Within 2 h after intraperitoneal injection of lipopolysaccharide, a product known to stimulate release of TNF, SAA preferably localized to ovarian epithelial cells and the thecal-interstitial layers compared to granulosal cell layers. Based on Gene Expression Omnibus (GEO) database, SAA1/2 and TNF were dominantly expressed in advanced grade ovarian cancer. Taken together, the accumulation of SAA1/2 in ovarian cancer could be mediated by TNF-induced NF-κB activation.
Keyword
MeSH Terms
-
Acute-Phase Proteins
Amyloid*
Cell Line
Delayed Diagnosis
Epithelial Cells
Gene Expression
Humans*
Immune System
Inflammation
Injections, Intraperitoneal
Mass Screening
Mortality
Necrosis
Ovarian Neoplasms
Serum Amyloid A Protein
Tumor Necrosis Factor-alpha
Acute-Phase Proteins
Amyloid
Serum Amyloid A Protein
Tumor Necrosis Factor-alpha
Figure
-

Figure 1 SAA1/2 is a dominant isoform in human ovarian tumor tissues. (A) Expression levels of SAA1/2 mRNA determined by real-time PCR in five different ovarian normal (N1-N5) and tumor tissues (T1-T5). (B) Expression levels of SAA4 mRNA in normal ovarian tissues and tumor specimens determined by real-time PCR. (C) Visual confirmation of SAA1/2 and SAA4 expression by RT-PCR. After isolating total RNA, RT-PCR and real-time PCR were carried out using primers for SAA1/2 and SAA4. GAPDH was used as a loading control.
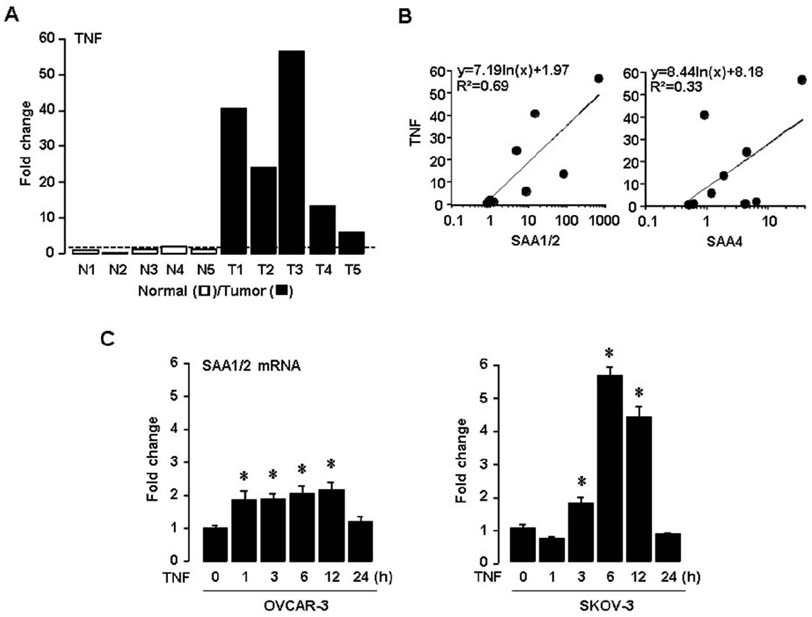
Figure 2 TNF enhances SAA1/2 expression in ovarian cancer. (A) Expression levels of TNF mRNA determined by real-time PCR in five different ovarian normal (N1-N5) and tumor tissues (T1-T5). (B) Correlation between mRNA levels of TNF and SAA isoforms. (C) Time-dependent effects of TNF on SAA1/2 mRNA by real-time PCR in ovarian cancer cell lines (OVCAR-3 and SKOV-3). Cells were incubated with TNF (10 ng/ml) for 0, 1, 3, 6, 12 and 24 hours. *Indicates significant increase (p≤0.05) compared to its own control as calculated by the paired Student's t test.
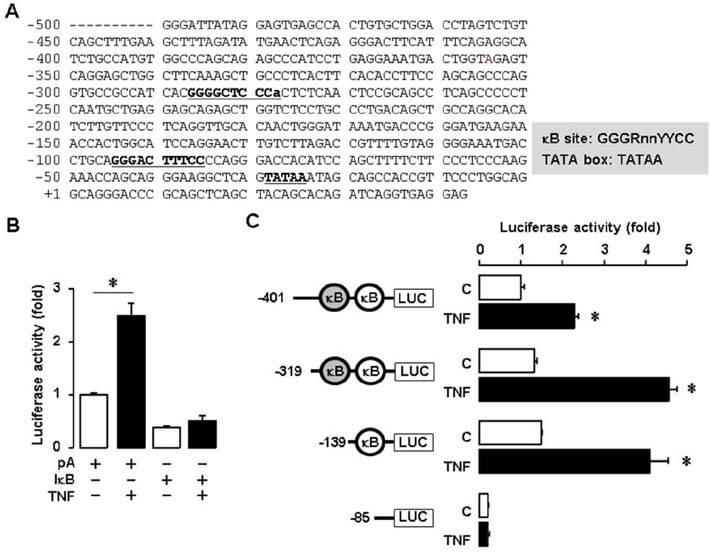
Figure 3 TNF increases SAA1 promoter activity via NF-κB signaling. (A) DNA sequences of the human SAA1 promoter. (B) Effect of TNF and IκB on luciferase activity of SAA1 promoter in SKOV-3 cells. Results were normalized to the protein level and represented as a fold increase compared to empty vector (pA) controls. (C) Effects of TNF on luciferase activity of the SAA1 promoter with NF-κB site deletions. After SKOV-3 cells were transfected with SAA1 luciferase vectors overnight, cells were treated with TNF (10 ng/ml) for 6 h, followed by luciferase assay. Results were normalized to the protein level and represented as a fold increase compared to non-treated control. Grey and white colors indicate NF-κB like (lower case for unmatched DNA sequence) and consensus sites, respectively. *Indicates significant increase (p≤0.05) compared to its own control as calculated by the paired Student's t test.
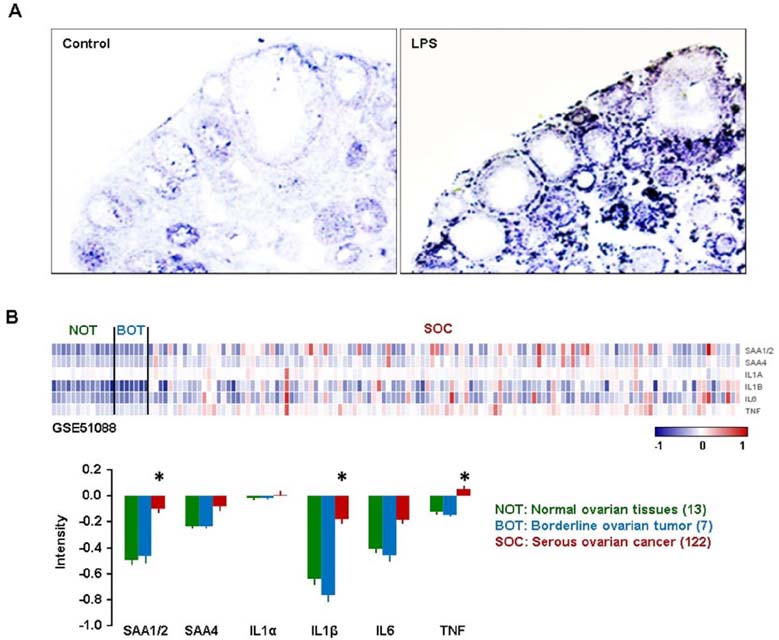
Figure 4 Ovarian localization of SAA after LPS treatment in mice, and GEO profiles of SAAs and proinflammatory cytokines in human ovarian tumor tissues. (A) LPS-induced SAA3 accumulation in ovarian epithelial and thecal-interstitial layers of mouse ovary. Six week-old mice were given vehicle (saline) or LPS (100 µg/mouse, ip). Ovaries were collected at 2 hrs after LPS treatment and ovarian localization of SAA3 mRNA was investigated by using in situ hybridization with a DIG-labeled RNA probe. Sense probe of SAA did not express any signal (data not shown). (B) GEO profiles of SAAs and proinflammatory cytokines in normal ovarian tissues (NOT), benign ovarian tumor (BOT) and serous ovarian cancer (SOC) based on the NCBI GEO database (GSE51088). Data values were expressed as the mean±SEM. *indicate significantly higher (p≤0.05) within groups as analyzed by Tukey's pairwise comparisons.
Reference
-
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016; 66:7–30.
Article2. Shih IM, Kurman RJ. Ovarian tumorigenesis: a proposed model based on morphological and molecular genetic analysis. Am J Pathol. 2004; 164:1511–1518.3. Kobel M, Kalloger SE, Boyd N, McKinney S, Mehl E, Palmer C, Leung S, Bowen NJ, Ionescu DN, Rajput A, Prentice LM, Miller D, Santos J, Swenerton K, Gilks CB, Huntsman D. Ovarian carcinoma subtypes are different diseases: implications for biomarker studies. PLoS Med. 2008; 5:e232.
Article4. Nowak M, Glowacka E, Szpakowski M, Szyllo K, Malinowski A, Kulig A, Tchorzewski H, Wilczynski J. Proinflammatory and immunosuppressive serum, ascites and cyst fluid cytokines in patients with early and advanced ovarian cancer and benign ovarian tumors. Neuro Endocrinol Lett. 2010; 31:375–383.5. Clendenen TV, Lundin E, Zeleniuch-Jacquotte A, Koenig KL, Berrino F, Lukanova A, Lokshin AE, Idahl A, Ohlson N, Hallmans G, Krogh V, Sieri S, Muti P, Marrangoni A, Nolen BM, Liu M, Shore RE, Arslan AA. Circulating inflammation markers and risk of epithelial ovarian cancer. Cancer Epidemiol Biomarkers Prev. 2011; 20:799–810.
Article6. Son DS, Kabir SM, Dong Y, Lee E, Adunyah SE. Characteristics of chemokine signatures elicited by EGF and TNF in ovarian cancer cells. J Inflamm (Lond). 2013; 10:25.
Article7. Ignacio RM, Kabir SM, Lee ES, Adunyah SE, Son DS. NF-kappaB-mediated CCL20 reigns dominantly in CXCR2-driven ovarian cancer progression. PLoS One. 2016; 11:e0164189.8. Dong YL, Kabir SM, Lee ES, Son DS. CXCR2-driven ovarian cancer progression involves upregulation of proinflammatory chemokines by potentiating NF-kappaB activation via EGFR-transactivated Akt signaling. PLoS One. 2013; 8:e83789.9. Dobrzycka B, Terlikowski SJ, Kowalczuk O, Kinalski M. Circulating levels of TNF-alpha and its soluble receptors in the plasma of patients with epithelial ovarian cancer. Eur Cytokine Netw. 2009; 20:131–134.
Article10. Maccio A, Madeddu C. Inflammation and ovarian cancer. Cytokine. 2012; 58:133–147.
Article11. Szlosarek PW, Grimshaw MJ, Kulbe H, Wilson JL, Wilbanks GD, Burke F, Balkwill FR. Expression and regulation of tumor necrosis factor alpha in normal and malignant ovarian epithelium. Mol Cancer Ther. 2006; 5:382–390.
Article12. Urieli-Shoval S, Linke RP, Matzner Y. Expression and function of serum amyloid A, a major acute-phase protein, in normal and disease states. Curr Opin Hematol. 2000; 7:64–69.
Article13. Targonska-Stepniak B, Majdan M. Serum amyloid A as a marker of persistent inflammation and an indicator of cardiovascular and renal involvement in patients with rheumatoid arthritis. Mediators Inflamm. 2014; 2014:793628.14. Thorn CF, Lu ZY, Whitehead AS. Regulation of the human acute phase serum amyloid A genes by tumour necrosis factor-alpha, interleukin-6 and glucocorticoids in hepatic and epithelial cell lines. Scand J Immunol. 2004; 59:152–158.
Article15. Urieli-Shoval S, Finci-Yeheskel Z, Dishon S, Galinsky D, Linke RP, Ariel I, Levin M, Ben-Shachar I, Prus D. Expression of serum amyloid a in human ovarian epithelial tumors: implication for a role in ovarian tumorigenesis. J Histochem Cytochem. 2010; 58:1015–1023.
Article16. Kilkenny C, Browne WJ, Cuthill IC, Emerson M, Altman DG. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS Biol. 2010; 8:e1000412.
Article17. Son DS, Roby KF. Interleukin-1alpha-induced chemokines in mouse granulosa cells: impact on keratinocyte chemoattractant chemokine, a CXC subfamily. Mol Endocrinol. 2006; 20:2999–3013.
Article18. Son DS, Terranova PF, Roby KF. Interaction of adenosine 3',5'-cyclic monophosphate and tumor necrosis factor-alpha on serum amyloid A3 expression in mouse granulosa cells: dependence on CCAAT-enhancing binding protein-beta isoform. Endocrinology. 2010; 151:3407–3419.
Article19. Perez-Llamas C, Lopez-Bigas N. Gitools: analysis and visualisation of genomic data using interactive heat-maps. PLoS One. 2011; 6:e19541.
Article20. Son DS, Roby KF, Terranova PF. Tumor necrosis factor-alpha induces serum amyloid A3 in mouse granulosa cells. Endocrinology. 2004; 145:2245–2252.
Article21. Hoesel B, Schmid JA. The complexity of NF-kappaB signaling in inflammation and cancer. Mol Cancer. 2013; 12:86.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Inhibitory mechanism on NF-kB transactivation by dexamethasone in pulmonary epithelial cells
- Effect of Transcription Factor Decoy for NF-κB on the TNF-α Induced Cytokine and ICAM-1 Expression in Cultured HaCaT cells
- Pro-inflammatory cytokine expression through NF-kappaB/IkappaB pathway in lung epithelial cells
- Expression of p53, c-myc, Transforming Growth Factor-alpha and -beta in Human Epithelial Ovarian Tumors
- Triptolide-induced Transrepression of IL-8 NF-kappaB in Lung Epithelial Cells
