Col1a1-cre mediated activation of beta-catenin leads to aberrant dento-alveolar complex formation
- Affiliations
-
- 1Cluster for Craniofacial Development and Regeneration Research, Institute of Oral Biosciences, Chonbuk National University School of Dentistry, Jeonju, Korea. oasis@jbnu.ac.kr
- 2Department of Pharmacology, Graduate School of Medicine, Kyoto University, Kyoto, Japan.
- KMID: 2046747
- DOI: http://doi.org/10.5115/acb.2012.45.3.193
Abstract
- Wnt/beta-catenin signaling plays a critical role in bone formation and regeneration. Dentin and cementum share many similarities with bone in their biochemical compositions and biomechanical properties. Whether Wnt/beta-catenin signaling is involved in the dento-alveolar complex formation is unknown. To understand the roles of Wnt/beta-catenin signaling in the dento-alveolar complex formation, we generated conditional beta-catenin activation mice through intercross of Catnb+/lox(ex3) mice with Col1a1-cre mice. In mutant mice, tooth formation and eruption was disturbed. Lower incisors and molars did not erupt. Bone formation was increased in the mandible but tooth formation was severely disturbed. Hypomineralized dentin was deposited in the crown but roots of molars were extremely short and distorted. In the odontoblasts of mutant molars, expression of dentin matrix proteins was obviously downregulated following the activation of beta-catenin whereas that of mineralization inhibitor was increased. Cementum and periodontal ligament were hypoplastic but periodontal space was narrow due to increased alveolar bone formation. While cementum matrix proteins were decreased, bone matrix proteins were increased in the cementum and alveolar bone of mutant mice. These results indicate that local activation of beta-catenin in the osteoblasts and odontoblasts leads to aberrant dento-alveolar complex formation. Therefore, appropriate inhibition of Wnt/beta-catenin signaling is important for the dento-alveolar complex formation.
MeSH Terms
Figure
-

Fig. 1 Targeted activation of β-catenin in the dentoalveolar complex. (A) In immunohistochemistry, β-catenin is localized in the Am, Od, and Ob of developing mouse dentoalveolar complex at P8. (B) β-galactosidase activities are also found in the Od and Ob of Col1a1-cre:R26R double transgenic mouse dentoalveolar complex at P8. (C) Col1a1-cre:Catnb+/lox(ex3) mutant mice exhibit severe growth retardation with short height and small body weights. (D) Differences in body weights between WT and MT mice are appeared in 3 weeks after birth and are clearer in the mice at 4 weeks-old. (E) Genotype analysis of mutant mice after intercross with Col1a1-cre and Catnb+/lox(ex3) mice. (F-I) Gross appearance of incisors in WT and MT mice at 4 weeks-old. Both of upper and lower incisors are normally erupted in WT mice, whereas upper incisors are erupted but lower incisors (white arrow) are not erupted in MT mice. (J, K) Molars of lower jaw (white arrowheads) are not erupted into oral cavity in MT mice in contrast to normally erupted in WT mice. Am, ameloblasts; Od, odontoblasts; Ob, osteoblasts; WT, wild type; MT, mutant. *P<0.05. Scale bar=200 µm (A, B).

Fig. 2 Disturbances in tooth formation and eruption failure of molars in Col1a1-cre:Catnb+/lox(ex3) mice. (A, B) In microradiography, general dimensions of craniofacial skeleton in MT mice are smaller than those in WT littermates. Particularly, mandible of MT is severely retarded and resulted in malocclusion. (C, D) In the mid-sagittal view, mandibular incisor is shorter and smaller than that of WT mice. (E, F) Molars of MT mice are impacted within mandible and root formation is impaired while those of WT mice are normally formed and erupted into the oral cavity. (G, H) In H&E-stained sagittal sections of the mandibles, bone deposition is remarkably increased and molars are not erupted in the MT mice. (I, J) In MT mice, β-catenin expression is upregulated in the odontoblasts and osteoblasts. MT, mutant; WT, wild type; H&E, hematoxylin and eosin. Scale bar=100 µm (G-J).
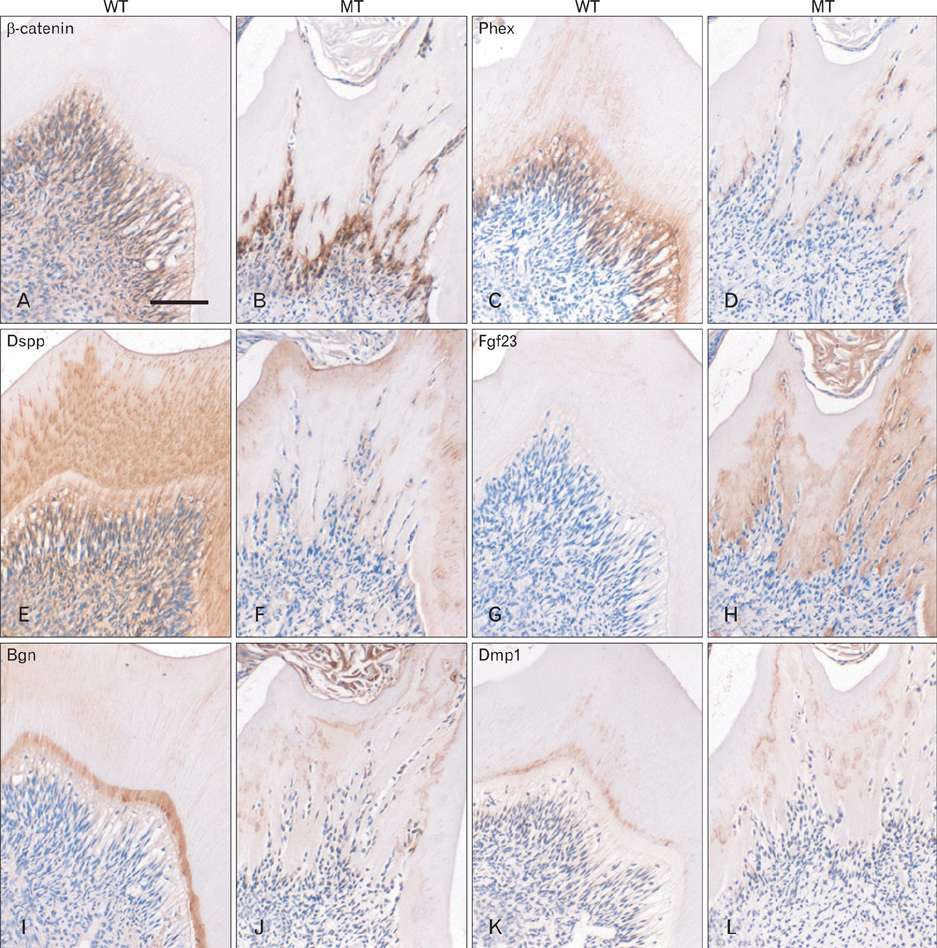
Fig. 3 Mineralization defects in dentin of Col1a1-cre:Catnb+/lox(ex3) mice. (A, B) Immunohistochemistry reveal that β-catenin is localized in the Od and pulp cells of mandibular first molar of WT mice, which is restrictedly increased in the odontoblasts of MT mice. (C, D) Phex is localized in the odontoblasts of WT mice, but which is clearly reduced in the odontoblasts of MT mice. (E, F) In WT mice, Dspp is localized in the dentin as well as odontoblasts, but it is obviously decreased in the predentin and odontoblasts of MT mice. Dspp remains only in thin mineralized dentin of MT mice. (G, H) Localization of Fgf23 is not found in the dentin and odontoblasts of WT mice but which is clearly observed in the predentin of MT mice. (I-L) Bgn and Dmp1 are mainly localized in the predentin and a part of dentin in WT mice, which also decreased in the predentin of MT mice. Od, odontoblasts; WT, wild type; MT, mutant; Phex, phosphate regulating endopeptidase homologue on the X chromosome; Dspp, dentin sialophosphoprotein; Fgf23, fibroblast growth factor-23; Bgn, biglycan; Dmp1, dentin matrix protein-1. Scale bar=100 µm (A-L).

Fig. 4 Deformities in the roots and periodontium of Col1a1-cre:Catnb+/lox(ex3) mice. (A, B) In contrast to WT mice, RD is thin and hypomineralized in MT mice. In addition, PS is narrower due to excessive formation of alveolar bone in MT mice. (C, D) In the RD of MT mice, Dspp is decreased in the dentin. (E-J) In WT mice, Bsp, Dmp1 and Fgf23 are specifically localized in the matrix of acellular cementum (arrows) and alveolar bone, which is decreased in the MT mice. (K, L) Osteopontin is localized in the cementum and periodontal ligaments of WT mice. It is also decreased in the cementum and periodontal ligaments of MT mice. WT, wild type; RD, root dentin; H&E, hematoxylin and eosin; MT, mutant; PS, periodontal space; Dspp, dentin sialophosphoprotein; Bsp, bone sialoprotein; Dmp1, dentin matrix protein-1; Fgf23, fibroblast growth factor-23; Opn, osteopontin. Scale bar=50 µm (A-L).
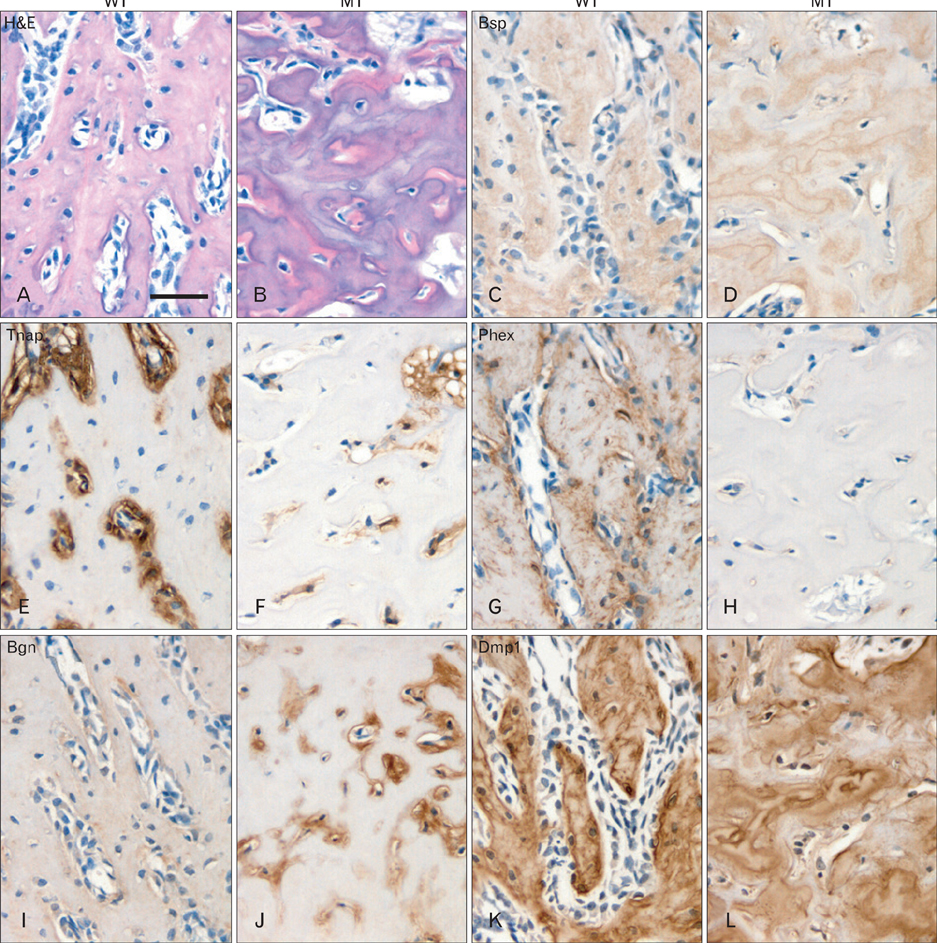
Fig. 5 Molecular changes related with increased bone mass in the mandibles of Col1a1-cre:Catnb+/lox(ex3) mice. (A, B) In the mandible of MT mice, bone mass is increased but number of osteoblasts are decreased to compare with those of WT mice. (C-F) Bsp expression is slightly increased with increase of bone mass, but Tnap is clearly decreased in the MT. (G-J) Bgn is significantly increased in the mandible of MT mice, while Phex is almost disappeared in MT mice. (K, L) Increased expression of Dmp1 is also observed in MT mice. WT, wild type; H&E, hamatoxylin and eosin; MT, mutant; Bsp, bone sialoprotein; Tnap, tissue-nonspecific alkaline phosphatase; Bgn, biglycan; Phex, phosphate regulating endopeptidase homologue on the X chromosome; Dmp1, dentin matrix protein-1. Scale bar=50 µm (A-L).
Reference
-
1. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004. 20:781–810.2. Reya T, Clevers H. Wnt signaling in stem cells and cancer. Nature. 2005. 434:843–850.3. Glass DA 2nd, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, Taketo MM, Long F, McMahon AP, Lang RA, Karsenty G. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell. 2005. 8:751–764.4. Holmen SL, Zylstra CR, Mukherjee A, Sigler RE, Faugere MC, Bouxsein ML, Deng L, Clemens TL, Williams BO. Essential role of beta-catenin in postnatal bone acquisition. J Biol Chem. 2005. 280:21162–21168.5. Gong Y, Slee RB, Fukai N, Rawadi G, Roman-Roman S, Reginato AM, Wang H, Cundy T, Glorieux FH, Lev D, Zacharin M, Oexle K, Marcelino J, Suwairi W, Heeger S, Sabatakos G, Apte S, Adkins WN, Allgrove J, Arslan-Kirchner M, Batch JA, Beighton P, Black GC, Boles RG, Boon LM, Borrone C, Brunner HG, Carle GF, Dallapiccola B, De Paepe A, Floege B, Halfhide ML, Hall B, Hennekam RC, Hirose T, Jans A, Jüppner H, Kim CA, Keppler-Noreuil K, Kohlschuetter A, LaCombe D, Lambert M, Lemyre E, Letteboer T, Peltonen L, Ramesar RS, Romanengo M, Somer H, Steichen-Gersdorf E, Steinmann B, Sullivan B, Superti-Furga A, Swoboda W, van den Boogaard MJ, Van Hul W, Vikkula M, Votruba M, Zabel B, Garcia T, Baron R, Olsen BR, Warman ML. Osteoporosis-Pseudoglioma Syndrome Collaborative Group. LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell. 2001. 107:513–523.6. Kato M, Patel MS, Levasseur R, Lobov I, Chang BH, Glass DA 2nd, Hartmann C, Li L, Hwang TH, Brayton CF, Lang RA, Karsenty G, Chan L. Cbfa1-independent decrease in osteoblast proliferation, osteopenia, and persistent embryonic eye vascularization in mice deficient in Lrp5, a Wnt coreceptor. J Cell Biol. 2002. 157:303–314.7. Boyden LM, Mao J, Belsky J, Mitzner L, Farhi A, Mitnick MA, Wu D, Insogna K, Lifton RP. High bone density due to a mutation in LDL-receptor-related protein 5. N Engl J Med. 2002. 346:1513–1521.8. Little RD, Carulli JP, Del Mastro RG, Dupuis J, Osborne M, Folz C, Manning SP, Swain PM, Zhao SC, Eustace B, Lappe MM, Spitzer L, Zweier S, Braunschweiger K, Benchekroun Y, Hu X, Adair R, Chee L, FitzGerald MG, Tulig C, Caruso A, Tzellas N, Bawa A, Franklin B, McGuire S, Nogues X, Gong G, Allen KM, Anisowicz A, Morales AJ, Lomedico PT, Recker SM, Van Eerdewegh P, Recker RR, Johnson ML. A mutation in the LDL receptor-related protein 5 gene results in the autosomal dominant high-bone-mass trait. Am J Hum Genet. 2002. 70:11–19.9. Liu F, Millar SE. Wnt/beta-catenin signaling in oral tissue development and disease. J Dent Res. 2010. 89:318–330.10. Liu F, Chu EY, Watt B, Zhang Y, Gallant NM, Andl T, Yang SH, Lu MM, Piccolo S, Schmidt-Ullrich R, Taketo MM, Morrisey EE, Atit R, Dlugosz AA, Millar SE. Wnt/beta-catenin signaling directs multiple stages of tooth morphogenesis. Dev Biol. 2008. 313:210–224.11. Chen J, Lan Y, Baek JA, Gao Y, Jiang R. Wnt/beta-catenin signaling plays an essential role in activation of odontogenic mesenchyme during early tooth development. Dev Biol. 2009. 334:174–185.12. Järvinen E, Salazar-Ciudad I, Birchmeier W, Taketo MM, Jernvall J, Thesleff I. Continuous tooth generation in mouse is induced by activated epithelial Wnt/beta-catenin signaling. Proc Natl Acad Sci U S A. 2006. 103:18627–18632.13. Kim TH, Lee JY, Baek JA, Lee JC, Yang X, Taketo MM, Jiang R, Cho ES. Constitutive stabilization of β-catenin in the dental mesenchyme leads to excessive dentin and cementum formation. Biochem Biophys Res Commun. 2011. 412:549–555.14. Harada N, Tamai Y, Ishikawa T, Sauer B, Takaku K, Oshima M, Taketo MM. Intestinal polyposis in mice with a dominant stable mutation of the beta-catenin gene. EMBO J. 1999. 18:5931–5942.15. Dacquin R, Starbuck M, Schinke T, Karsenty G. Mouse alpha1 (I)-collagen promoter is the best known promoter to drive efficient Cre recombinase expression in osteoblast. Dev Dyn. 2002. 224:245–251.16. Tiffee JC, Xing L, Nilsson S, Boyce BF. Dental abnormalities associated with failure of tooth eruption in src knockout and op/op mice. Calcif Tissue Int. 1999. 65:53–58.17. Kong YY, Yoshida H, Sarosi I, Tan HL, Timms E, Capparelli C, Morony S, Oliveira-dos-Santos AJ, Van G, Itie A, Khoo W, Wakeham A, Dunstan CR, Lacey DL, Mak TW, Boyle WJ, Penninger JM. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature. 1999. 397:315–323.18. Li J, Sarosi I, Yan XQ, Morony S, Capparelli C, Tan HL, McCabe S, Elliott R, Scully S, Van G, Kaufman S, Juan SC, Sun Y, Tarpley J, Martin L, Christensen K, McCabe J, Kostenuik P, Hsu H, Fletcher F, Dunstan CR, Lacey DL, Boyle WJ. RANK is the intrinsic hematopoietic cell surface receptor that controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc Natl Acad Sci U S A. 2000. 97:1566–1571.19. Kim N, Odgren PR, Kim DK, Marks SC Jr, Choi Y. Diverse roles of the tumor necrosis factor family member TRANCE in skeletal physiology revealed by TRANCE deficiency and partial rescue by a lymphocyte-expressed TRANCE transgene. Proc Natl Acad Sci U S A. 2000. 97:10905–10910.20. Lu X, Rios HF, Jiang B, Xing L, Kadlcek R, Greenfield EM, Luo G, Feng JQ. A new osteopetrosis mutant mouse strain (ntl) with odontoma-like proliferations and lack of tooth roots. Eur J Oral Sci. 2009. 117:625–635.21. Yamashiro T, Tummers M, Thesleff I. Expression of bone morphogenetic proteins and Msx genes during root formation. J Dent Res. 2003. 82:172–176.22. Nakatomi M, Morita I, Eto K, Ota MS. Sonic hedgehog signaling is important in tooth root development. J Dent Res. 2006. 85:427–431.23. Steele-Perkins G, Butz KG, Lyons GE, Zeichner-David M, Kim HJ, Cho MI, Gronostajski RM. Essential role for NFI-C/CTF transcription-replication factor in tooth root development. Mol Cell Biol. 2003. 23:1075–1084.24. Gao Y, Yang G, Weng T, Du J, Wang X, Zhou J, Wang S, Yang X. Disruption of Smad4 in odontoblasts causes multiple keratocystic odontogenic tumors and tooth malformation in mice. Mol Cell Biol. 2009. 29:5941–5951.25. Nemoto E, Koshikawa Y, Kanaya S, Tsuchiya M, Tamura M, Somerman MJ, Shimauchi H. Wnt signaling inhibits cementoblast differentiation and promotes proliferation. Bone. 2009. 44:805–812.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Role of E-cadherin/Beta-catenin Complex and Cyclin D1 in Head and Neck Squamous Cell Carcinoma
- Expression of E-cadherin/beta-catenin Complex in Uterine Cervix Cancer
- Euodia sutchuenensis Dode extract stimulates osteoblast differentiation via Wnt/beta-catenin pathway activation
- Expression of beta-Catenin, c-Myc, and Cyclin D1 in Pulmonary Adenocarcinomas
- Gene Alterations by Osteoblast-specific Transcription Factor Osterix in Postnatal Bone Formation
