LPS-Induced Migration of Peritoneal B-1 Cells is Associated with Upregulation of CXCR4 and Increased Migratory Sensitivity to CXCL12
- Affiliations
-
- 1Division of Pathology, Department of Molecular Cell Biology and Samsung Biomedical Research Institute, Sungkyunkwan University School of Medicine, Suwon, Korea. tjkim@skku.edu
- KMID: 1792975
- DOI: http://doi.org/10.3346/jkms.2012.27.1.27
Abstract
- B-1 cells, which constitute a predominant lymphocyte subset in serosal cavities and produce most of natural antibodies, are subdivided into the CD5+ B-1a and CD5- B-1b cell subpopulations, but the differential roles of B-1a and B-1b cells are not well understood. We report that B-1a cells preferentially migrate out of the peritoneal cavity and upregulate the expression of CXCR4 with heightened sensitivity to CXCL12 and CXCL13 upon LPS treatment compared to B-1b and B-2 cells. Whereas B-1a cells were slightly more abundant than B-1b and B-2 cells in the homeostatic condition, the number of B-1a cells preferentially decreased 48 hr after LPS treatment. The decrease in the peritoneal B-1a cell number was accompanied with increased migration of B-1a cells toward CXCL-12 and CXCL-13 in in vitro transmigration assay using peritoneal B cells from LPS treated mice. The expression level of CXCR4, but not of CXCR5, was also more prominently increased in B-1a cells upon LPS stimulation. LPS-stimulated B-1a cells did not accumulate in omental milky spots in contrast to B-2 cells. These results suggest that B-1a cells actively migrate out of the peritoneal cavity through the regulation of the migratory responsiveness to chemokines and actively participate in systemic immune responses.
Keyword
MeSH Terms
-
Adjuvants, Immunologic/pharmacology
Animals
B-Lymphocytes/cytology/*drug effects/immunology
Cell Movement
Cells, Cultured
Chemokine CXCL12/metabolism/*pharmacology
Chemokine CXCL13/metabolism/pharmacology
Lipopolysaccharides/*pharmacology
Mice
Mice, Inbred C57BL
Peritoneal Cavity/cytology
Receptors, CXCR4/*metabolism
Up-Regulation
Figure
-
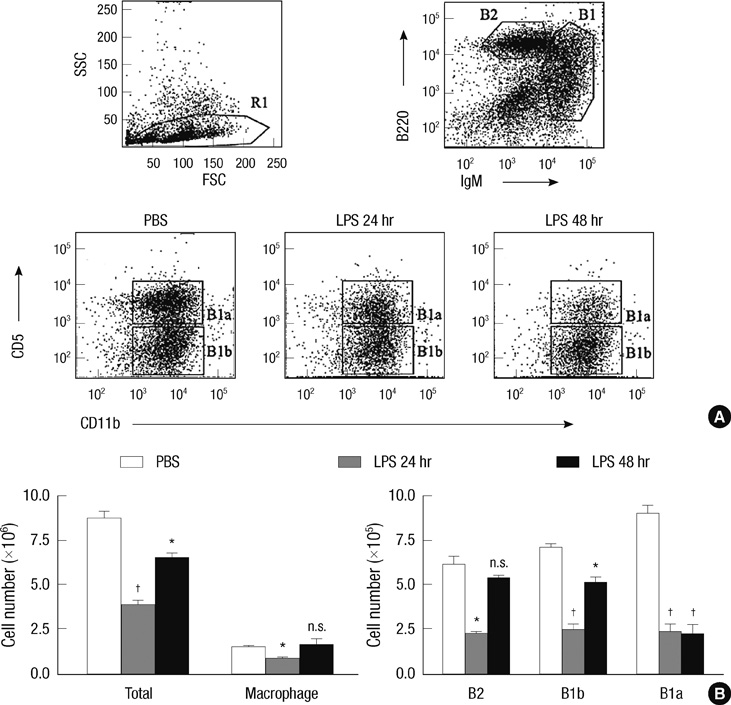
Fig. 1 LPS induces the preferential egress of peritoneal B-1a cells. (A) Peritoneal cells obtained from 10 week-old C57BL/6 mice at the indicated time after intraperitioneal injection of LPS were stained with fluorescence labeled anti-IgM, -B220, -CD11b, and -CD5 and were analyzed by flow cytometry. In all cases, doublets were excluded by forward scatter area vs height histogram. B-2 cells were gated based on B220high and IgMlow expression and B-1 cells were gated based on and IgMhigh expression. (B) At the indicated times after LPS injection, the absolute numbers of indicated cells in the peritoneal cavity were calculated from the total cell number and flow cytometric data. Representative flow cytometric data from three mice of each treatment group were shown in (A). Statistical analysis was performed by Student's t test in comparison with the numbers of PBS treatment group. n.s., differences not statistically significant. *P = 0.01-0.05; †P = 0.001-0.01.
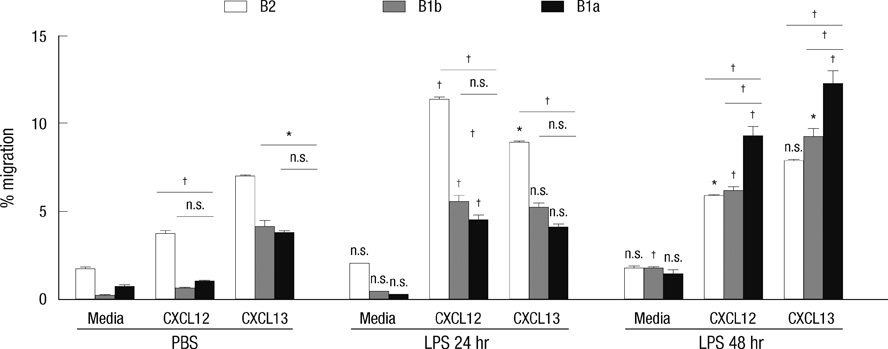
Fig. 2 Heightened migratory response of peritoneal B cells obtained from LPS-treated mice in response to CXCL12 or CXCL13. In vitro transwell migration assay was performed with peritoneal B cells isolated from 10 week-old C57BL/6 mice at the indicated time after intraperitioneal injection of LPS. Peritoneal B cells were added to the upper chamber of a transwell plate in the presence of CXCL12 or CXCL13 in the lower chamber. Two hour later, the numbers of B-1a, B-1b, and B-2 cells in the lower chambers were counted. Results represent the percentage of the cell number in the lower chamber over the input number in response to medium alone, CXCL12, or CXCL13. Data were collected from three mice each experimental group. Statistical analyses were performed by Student's t test for comparison between groups of treatment with PBS and LPS (24 and 48 hr) (above each bar) and also between B-1a cells and B-2 cells or B-1b cells (above each drawn line). n.s., differences not statistically significant. *P = 0.01-0.05; †P = 0.001-0.01.
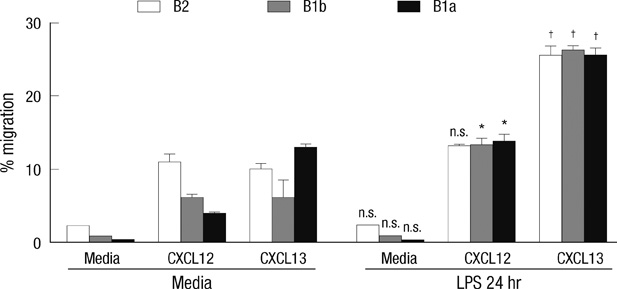
Fig. 3 Increased chemotactic response of LPS-stimulated peritoneal B cells in response to CXCL12 or CXCL13. In vitro migration assay was performed with purified peritoneal B cells stimulated for 24 hr with or without LPS in vitro. Peritoneal B cells were added to the upper chamber of transwell plate in the presence of CXCL12 or CXCL13 in the lower chamber. 2 hr later, the numbers of B-1a, B-1b, and B-2 cells in the lower chambers were counted. Results represent the percentage of the cell number in the lower chamber over the input number in response to medium alone, CXCL12, or CXCL13. Statistical analyses from three mice of each experimental group were performed by Student's t test in comparison with the group of media treatment. n.s., differences not statistically significant. *P = 0.01-0.05; †P = 0.001-0.01.

Fig. 4 Time-course expression of CXCR4 and CXCR5 in peritoneal B cell subsets upon LPS treatment. (A) Peritoneal cells isolated from 10 week-old C57BL/6 mice at the indicated time after intraperitioneal injection of LPS were stained with fluorescence labeled anti-IgM, -CD11b, -CD5, -CXCR4, and -CXCR5 and and were analyzed by flow cytometry. Histograms show mean fluorescence intensities (MFI) of surface CXCR4 (upper panel) and CXCR5 (lower panel). (B) Kinetics of the time-dependent expression of surface CXCR4 (left) or CXCR5 (right) on given peritoneal B cell subsets from C57BL/6 mice treated with LPS for given time duration.

Fig. 5 CXCR4, but not CXCR5, is differentially regulated in peritoneal B cell subsets upon LPS stimulation in vitro. (A) Purified peritoneal B cells were incubated for 24 hr with or without LPS and then were stained with fluorescence labeled anti-IgM, -CD11b, -CD5, -CXCR4, and -CXCR5 and were analyzed by flow cytometry. (B) Representative flow cytometric histograms of B cells are shown from three independent experiments. Numbers within the histograms are mean fluorescence intensities of surface CXCR4 or CXCR5 in given B cell subsets that are identified in (A).
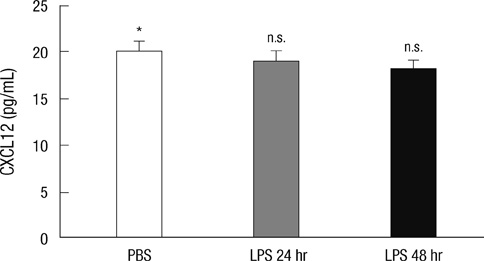
Fig. 6 The concentration of CXCL12 in the peritoneal fluids from LPS-injected mice. Peritoneal fluids isolated from 10 week-old C57BL/6 mice at the indicated time after intraperitioneal injection of LPS were measured for the CXCL12 concentrations by ELISA. Statistical analysis from three mice of each group was performed by Student's t test in comparison with the concentration of PBS treatment group. n.s., differences not statistically significant.
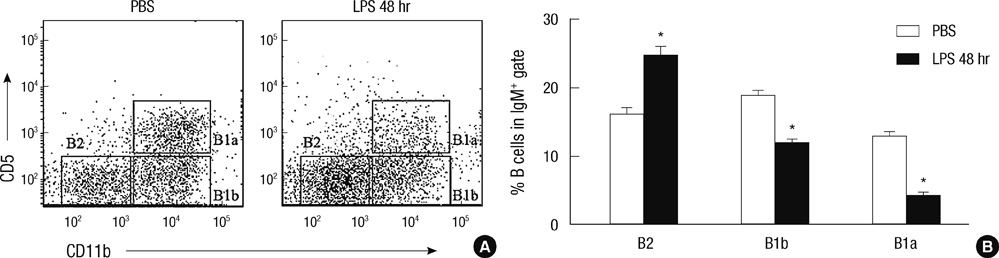
Fig. 7 B-1a cells do not migrate into the omental milky spots upon the LPS treatment in contrast to B-2 cells. (A) Omental cells obtained from 10 week-old C57BL/6 mice at 48 hr after intraperitioneal injection of LPS were stained with fluorescence labeled anti-IgM, -CD11b, and C-D5 and were analyzed by flow cytometry. (B) Percentages of given B cell subsets among total IgM+ B cells from the omental milky spots 48 hr after i.p. injection of LPS or PBS. Statistical analysis from three mice of each experimental group was performed by Student's t test in comparison with the percentage of PBS treatment group. *P = 0.01-0.05.
Cited by 2 articles
-
Expansion and Sub-Classification of T Cell-Dependent Antibody Responses to Encompass the Role of Innate-Like T Cells in Antibody Responses
Chanho Park, Tae Jin Kim
Immune Netw. 2018;18(5):. doi: 10.4110/in.2018.18.e34.The Role of B Cells in Transplantation Rejection
Tae Jin Kim
J Korean Soc Transplant. 2018;32(1):1-6. doi: 10.4285/jkstn.2018.32.1.1.
Reference
-
1. Montecino-Rodriguez E, Dorshkind K. New perspectives in B-1 B cell development and function. Trends Immunol. 2006. 27:428–433.2. Hardy RR. B-1 B cells: development, selection, natural autoantibody and leukemia. Curr Opin Immunol. 2006. 18:547–555.3. Montecino-Rodriguez E, Leathers H, Dorshkind K. Identification of a B-1 B cell-specified progenitor. Nat Immunol. 2006. 7:293–301.4. Alugupalli KR, Leong JM, Woodland RT, Muramatsu M, Honjo T, Gerstein RM. B1b lymphocytes confer T cell-independent long-lasting immunity. Immunity. 2004. 21:379–390.5. Szczepanik M, Akahira-Azuma M, Bryniarski K, Tsuji RF, Kawikova I, Ptak W, Kiener C, Campos RA, Askenase PW. B-1 B cells mediate required early T cell recruitment to elicit protein-induced delayed-type hypersensitivity. J Immunol. 2003. 171:6225–6235.6. Nogueira-Martins MF, Mariano M. B-1 cell participation in T-cell-mediated alloimmune response. Immunobiology. 2010. 215:264–274.7. Wardemann H, Boehm T, Dear N, Carsetti R. B-1a B cells that link the innate and adaptive immune responses are lacking in the absence of the spleen. J Exp Med. 2002. 195:771–780.8. Baggiolini M. Chemokines and leukocyte traffic. Nature. 1998. 392:565–568.9. Sallusto F, Mackay CR. Chemoattractants and their receptors in homeostasis and inflammation. Curr Opin Immunol. 2004. 16:724–731.10. Förster R, Mattis AE, Kremmer E, Wolf E, Brem G, Lipp M. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell. 1996. 87:1037–1047.11. Bowman EP, Campbell JJ, Soler D, Dong Z, Manlongat N, Picarella D, Hardy RR, Butcher EC. Developmental switches in chemokine response profiles during B cell differentiation and maturation. J Exp Med. 2000. 191:1303–1318.12. Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S, Kitamura Y, Yoshida N, Kikutani H, Kishimoto T. Defects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature. 1996. 382:635–638.13. Ma Q, Jones D, Borghesani PR, Segal RA, Nagasawa T, Kishimoto T, Bronson RT, Springer TA. Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice. Proc Natl Acad Sci U S A. 1998. 95:9448–9453.14. Palmesino E, Moepps B, Gierschik P, Thelen M. Differences in CXCR4-mediated signaling in B cells. Immunobiology. 2006. 211:377–389.15. Foussat A, Balabanian K, Amara A, Bouchet-Delbos L, Durand-Gasselin I, Baleux F, Couderc J, Galanaud P, Emilie D. Production of stromal cell-derived factor 1 by mesothelial cells and effects of this chemokine on peritoneal B lymphocytes. Eur J Immunol. 2001. 31:350–359.16. Yun HJ, Jo DY. Production of stromal cell-derived factor-1 (SDF-1)and expression of CXCR4 in human bone marrow endothelial cells. J Korean Med Sci. 2003. 18:679–685.17. Rangel-Moreno J, Moyron-Quiroz JE, Carragher DM, Kusser K, Hartson L, Moquin A, Randall TD. Omental milky spots develop in the absence of lymphoid tissue-inducer cells and support B and T cell responses to peritoneal antigens. Immunity. 2009. 30:731–743.18. Ansel KM, Ngo VN, Hyman PL, Luther SA, Förster R, Sedgwick JD, Browning JL, Lipp M, Cyster JG. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature. 2000. 406:309–314.19. Allen CD, Ansel KM, Low C, Lesley R, Tamamura H, Fujii N, Cyster JG. Germinal center dark and light zone organization is mediated by CXCR4 and CXCR5. Nat Immunol. 2004. 5:943–952.20. Caron G, Le Gallou S, Lamy T, Tarte K, Fest T. CXCR4 expression functionally discriminates centroblasts versus centrocytes within human germinal center B cells. J Immunol. 2009. 182:7595–7602.21. Ansel KM, Harris RB, Cyster JG. CXCL13 is required for B1 cell homing, natural antibody production, and body cavity immunity. Immunity. 2002. 16:67–76.22. Le Y, Zhu BM, Harley B, Park SY, Kobayashi T, Manis JP, Luo HR, Yoshimura A, Hennighausen L, Silberstein LE. SOCS3 protein developmentally regulates the chemokine receptor CXCR4-FAK signaling pathway during B lymphopoiesis. Immunity. 2007. 27:811–823.23. Nakashima H, Hamaguchi Y, Watanabe R, Ishiura N, Kuwano Y, Okochi H, Takahashi Y, Tamaki K, Sato S, Tedder TF, Fujimoto M. CD22 expression mediates the regulatory functions of peritoneal B-1a cells during the remission phase of contact hypersensitivity reactions. J Immunol. 2010. 184:4637–4645.24. Sugiyama T, Kohara H, Noda M, Nagasawa T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity. 2006. 25:977–988.25. Tarlinton D, Radbruch A, Hiepe F, Dörner T. Plasma cell differentiation and survival. Curr Opin Immunol. 2008. 20:162–169.26. Glodek AM, Honczarenko M, Le Y, Campbell JJ, Silberstein LE. Sustained activation of cell adhesion is a differentially regulated process in B lymphopoiesis. J Exp Med. 2003. 197:461–473.27. Baumgarth N. The double life of a B-1 cell: self-reactivity selects for protective effector functions. Nat Rev Immunol. 2011. 11:34–46.28. Chumley MJ, Dal Porto JM, Cambier JC. The unique antigen receptor signaling phenotype of B-1 cells is influenced by locale but induced by antigen. J Immunol. 2002. 169:1735–1743.29. Krist LF, Eestermans IL, Steenbergen JJ, Hoefsmit EC, Cuesta MA, Meyer S, Beelen RH. Cellular composition of milky spots in the human greater omentum: an immunochemical and ultrastructural study. Anat Rec. 1995. 241:163–174.30. Carlow DA, Gold MR, Ziltener HJ. Lymphocytes in the peritoneum home to the omentum and are activated by resident dendritic cells. J Immunol. 2009. 183:1155–1165.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Extracellular Hsp70 Is Involved in the CXCL12/CXCR4 Pathway in Primary Human Nasal Epithelial Cells: A Preliminary Study
- Targeting the CXCL12/CXCR4 axis in acute myeloid leukemia: from bench to bedside
- Isoorientin Suppresses Invasion of Breast and Colon Cancer Cells by Inhibition of CXC Chemokine Receptor 4 Expression
- The Role of SDF-1α-CXCR4/CXCR7 in Migration of Human Periodontal Ligament Stem Cells
- Expression and functional roles of the chemokine receptor CXCR7 in acute myeloid leukemia cells
