Cryptotanshinone promotes brown fat activity by AMPK activation to inhibit obesity
- Affiliations
-
- 1Department of Endocrinology, Affiliated Xiaoshan Hospital, Hangzhou Normal University, Hangzhou, Zhejiang 311202, China
- KMID: 2558487
- DOI: http://doi.org/10.4162/nrp.2024.18.4.479
Abstract
- BACKGROUND/OBJECTIVES
Activating brown adipose tissue (BAT) and browning of white adipose tissue (WAT) can protect against obesity and obesity-related metabolic conditions. Cryptotanshinone (CT) regulates lipid metabolism and significantly ameliorates insulin resistance. Adenosine-5'-monophosphate (AMP)-activated protein kinase (AMPK), a receptor for cellular energy metabolism, is believed to regulate brown fat activity in humans.
MATERIALS/METHODS
The in vivo study included high-fat-fed obese mice administered orally 200/400 mg/kg/d CT. They were evaluated through weight measurement, the intraperitoneal glucose tolerance test (IPGTT), intraperitoneal insulin tolerance test (IPITT), cold stimulation test, serum lipid (total cholesterol, triglycerides, and low-density lipoprotein) measurement, hematoxylin and eosin staining, and immunohistochemistry. Furthermore, the in vitro study investigated primary adipose mesenchymal stem cells (MSCs) with incubation of CT and AMPK agonists (acadesine)/inhibitor (Compound C). Cells were evaluated using Oil Red O staining, Alizarin red staining, flow cytometry, and immunofluorescence staining to identify and observe the osteogenic versus adipogenic differentiation. Quantitative real-time polymerase chain reaction and the Western blot were used to observe related gene expression.
RESULTS
In the diet-induced obesity mouse model mice CT suppressed body weight, food intake, glucose levels in the IPGTT and IPTT, serum lipids, the volume of adipose tissue, and increased thermogenesis, uncoupling protein 1, and the AMPK pathway expression. In the in vitro study, CT prevented the formation of lipid droplets from MSCs while activating brown genes and the AMPK pathway. AMPK activator enhanced CT’s effects, while the AMPK inhibitor reversed the effects of CT.
CONCLUSION
CT promotes adipose tissue browning to increase body thermogenesis and reduce obesity by activating the AMPK pathway. This study provides an experimental foundation for the use of CT in obesity treatment.
Keyword
Figure
-
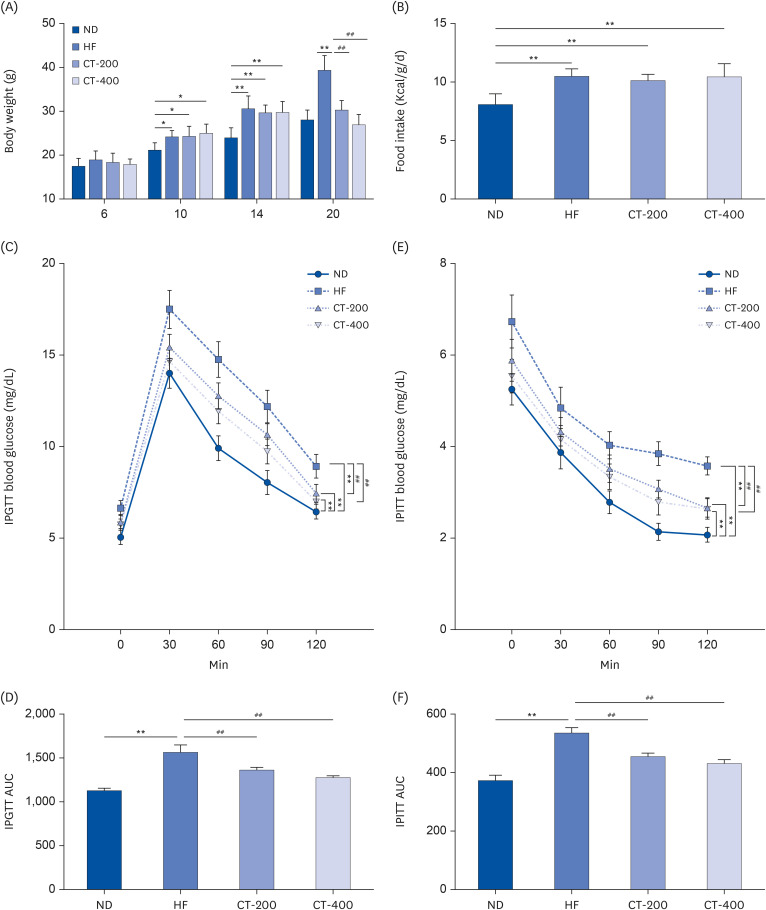
Fig. 1 CT reduces body weight and improves glucose intolerance and insulin resistance in mice. (A) Mice body weight and (B) food intake were examined (n = 6). (C) IPGTT was used to examine the effect of CT on glucose tolerance in mice on HF group. (D) AUC was calculated for IPGTT (n = 6). (E) IPITT was used to evaluate the effect of CT on insulin tolerance in mice on HF group (n = 6). (F) AUC was calculated for IPITT (n = 6). Data from A, B, D and F were analyzed by 1-way ANOVA. Data from C and E were analyzed by 2-way ANOVA.Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; IPGTT, intraperitoneal glucose tolerance test; AUC, area under the curve; IPITT, intraperitoneal insulin tolerance test; ANOVA, analysis of variance.*P < 0.05, **P < 0.01 vs. ND; ##P < 0.01 vs. HF.
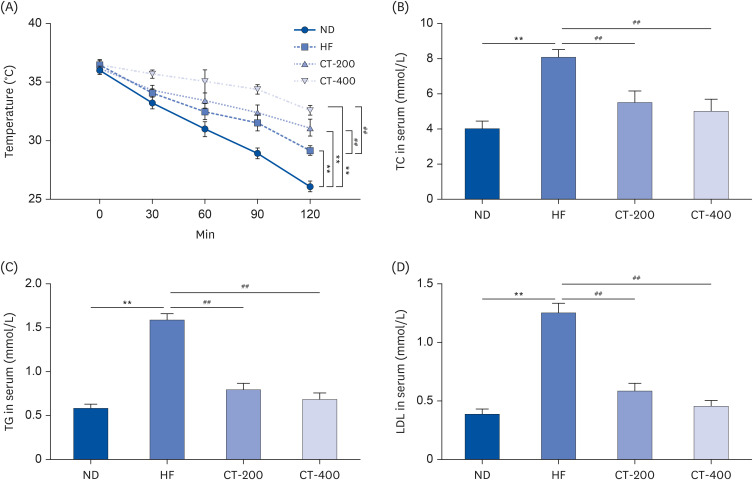
Fig. 2 CT increases adaptive thermogenesis and suppresses dyslipidemia. (A) The cold stimulation test was used to examine the regulation of body temperature in mice by CT and the data were analyzed by 2-way ANOVA (n = 6). (B-D) Serum TC, TG, and LDL-C levels were measured in each group of mice, and the data were analyzed by 1-way ANOVA (n = 6).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; ANOVA, analysis of variance; TC, total cholesterol; TG, triglycerides; LDL-C, low-density lipoprotein cholesterol; ANOVA, analysis of variance.**P < 0.01 vs. ND; ##P < 0.01 vs. HF.
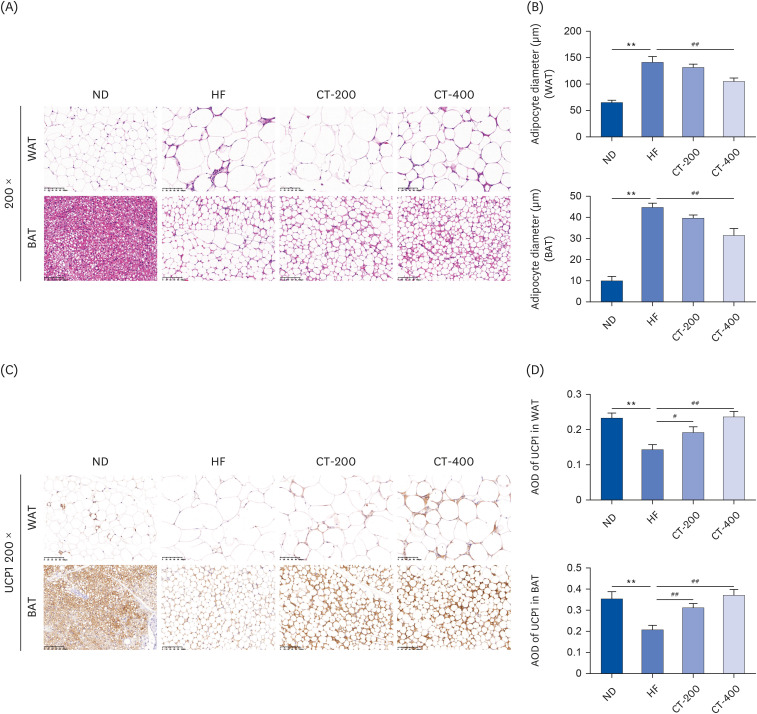
Fig. 3 CT improved adipose tissue morphology and promoted UCP1 levels. (A) Morphological effects of CT on WAT and BAT in mice as observed by H&E staining (magnification, 200×). (B) Average adipocyte diameter for quantitative analysis. (C, D) Immunohistochemistry was used to detect the effect of CT on UCP1 expression levels in WAT and BAT (magnification, 200×), and the data were analyzed by 1-way ANOVA (n = 3).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; UCP1, uncoupling protein 1; WAT, white adipose tissue; BAT, brown adipose tissue; H&E, hematoxylin and eosin; ANOVA, analysis of variance.**P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
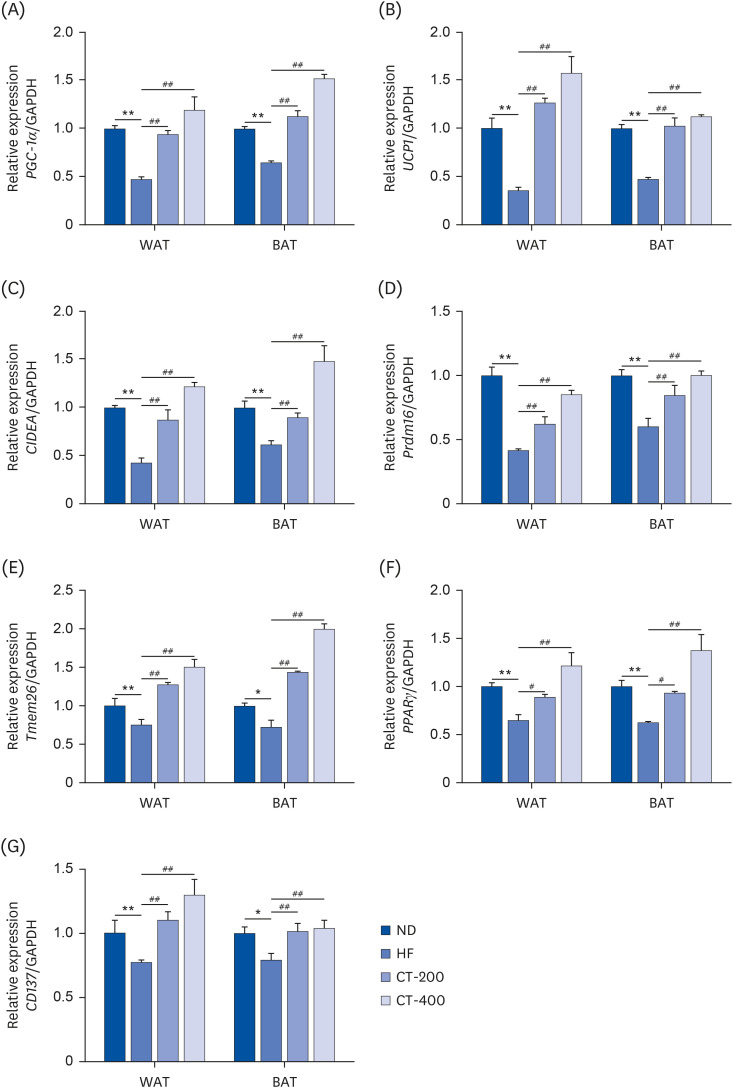
Fig. 4 CT promotes brown gene expression in adipose tissue. qRT-PCR was used to examine the effect of CT on the brown genes (A) PGC-1α, (B) UCP1, (C) CIDEA, (D) Prdm16, (E) Tmem26, (F) PPARγ, and (G) CD137 mRNA levels in the WAT and BAT tissues. Data were analyzed by one-way ANOVA (n = 3).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; qRT-PCR, quantitative real-time polymerase chain reaction; PGC-1α, proliferator-activated receptor-gamma coactivator-1α; UCP1, uncoupling protein 1; CIDEA, cell death inducing DFFA like effector A; Prdm16, prdomain-containing 16; Tmem26, transmembrane protein 26; PPARγ, peroxisome proliferator-activated receptor γ; CD, cluster of differentiation; WAT, white adipose tissue; BAT, brown adipose tissue; ANOVA, analysis of variance; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
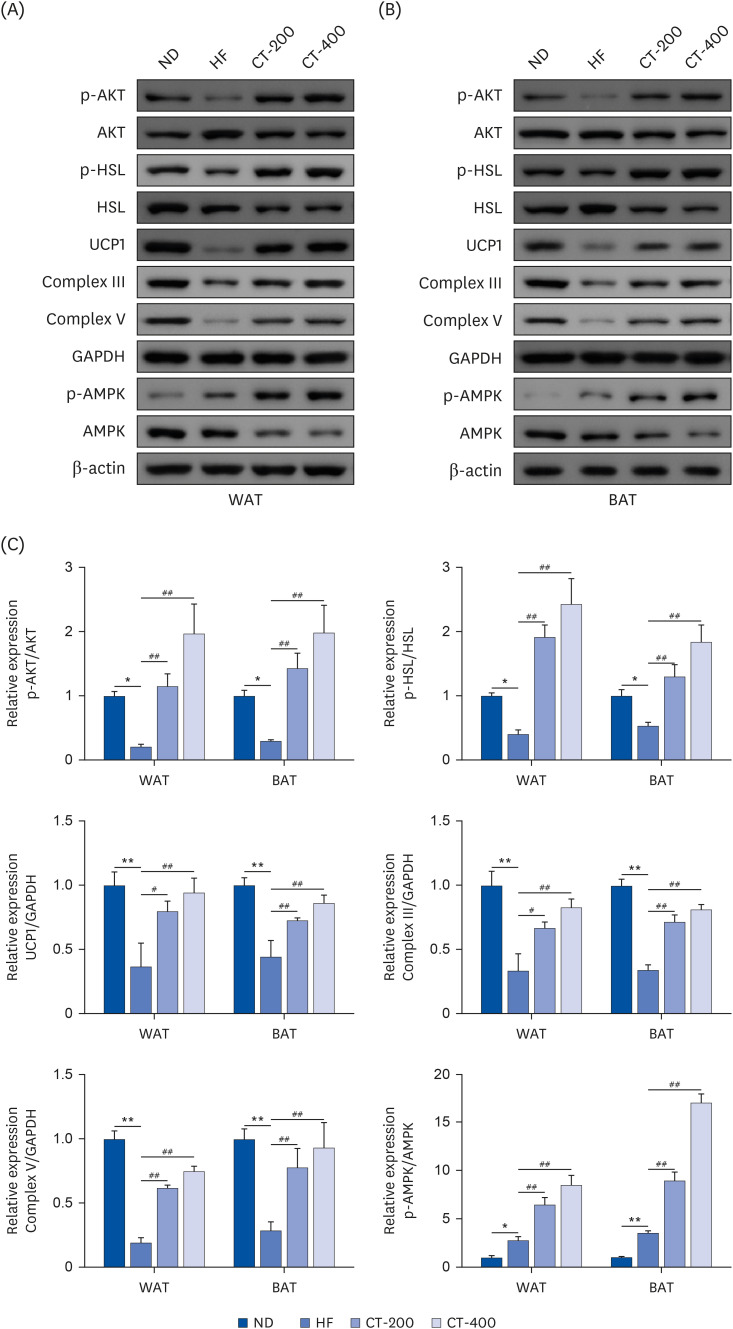
Fig. 5 CT promotes brown gene protein expression and improves insulin sensitivity and lipolysis in adipose tissue. Western blot was used to detect the effect of CT on protein expression levels of p-AKT/AKT, p-HSL/HSL, UCP1, Complex III, Complex V, and p-AMPK/AMPK in (A) WAT and (B) BAT. (C) qRT-PCR was used to examine the effect of CT on protein expression levels above. Data were analyzed by one-way ANOVA (n = 3).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; AKT, protein kinase B; HSL, hormone-sensitive triglyceride lipase; UCP1, uncoupling protein 1; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; WAT, white adipose tissue; BAT, brown adipose tissue; ANOVA, analysis of variance.*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
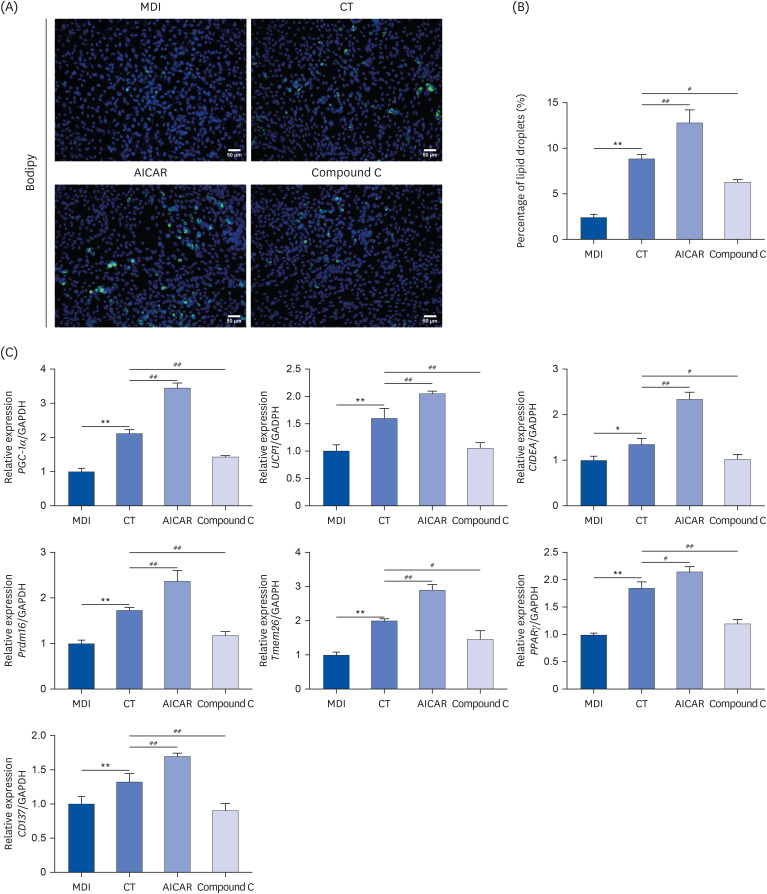
Fig. 6 CT inhibits lipid droplet formation and mRNA levels of brown genes in MSCs through activation of AMPK. (A, B) Immunofluorescence assays were used to detect lipid droplet formation in MSCs (magnification, 200×). (C) qRT-PCR was used to examine the effect of CT on the brown genes PGC-1α, UCP1, CIDEA, Prdm16, Tmem26, PPARγ, CD137 mRNA levels in MSCs. Data were analyzed by one-way ANOVA (n = 3).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; MSC, mesenchymal stem cell; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; qRT-PCR, quantitative real-time polymerase chain reaction; PGC-1α, proliferator-activated receptor-gamma coactivator-1α; UCP1, uncoupling protein 1; CIDEA, cell death inducing DFFA like effector A; Prdm16, prdomain-containing 16; Tmem26, transmembrane protein 26.PPARγ, peroxisome proliferator-activated receptor γ; CD, cluster of differentiation; ANOVA, analysis of variance.*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
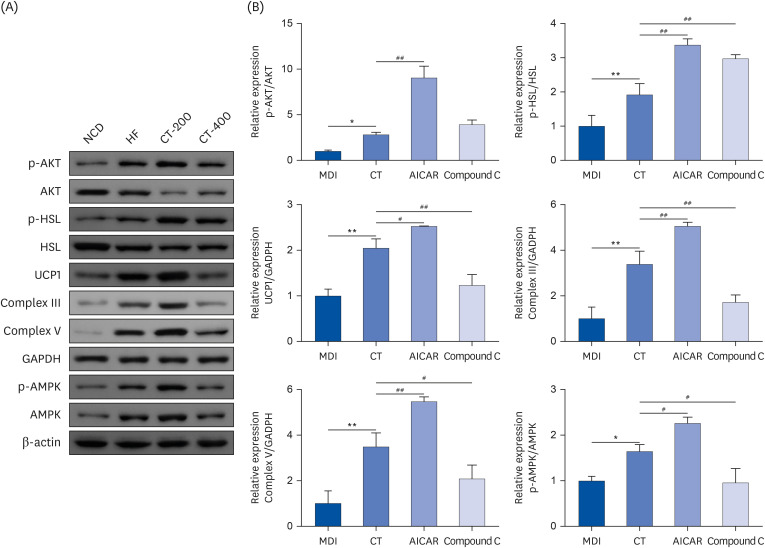
Fig. 7 CT promotes brown gene expression and improves insulin sensitivity and lipolysis in adipose tissue through activation of AMPK. (A, B) Western blot was used to detect the effect of CT on protein expression levels of p-AKT/AKT, p-HSL/HSL, UCP1, Complex III, Complex V, p-AMPK/AMPK in MSCs. Data were analyzed by one-way ANOVA (n = 3).Mice were randomly divided into 4 groups: ND, normal diet; HF, high-fat diet; CT-200, received 200 mg/kg/d CT; and CT-400, received 400 mg/kg/d CT (CT groups received via gavage for 6 weeks, respectively).CT, Cryptotanshinone; AMPK, adenosine-5'-monophosphate (AMP)-activated protein kinase; AKT, protein kinase B; HSL, hormone-sensitive triglyceride lipase; UCP1, uncoupling protein 1; MSC, mesenchymal stem cell; ANOVA, analysis of variance.*P < 0.05, **P < 0.01 vs. ND; #P < 0.05, ##P < 0.01 vs. HF.
Reference
-
1. Martínez-Vizcaíno V, Álvarez-Bueno C, Cavero-Redondo I. Diet in the management of weight loss. Nutrients. 2021; 13:1306. PMID: 33920924.
Article2. Diamantis DV, Karatzi K, Kantaras P, Liatis S, Iotova V, Bazdraska Y, Tankova T, Cardon G, Wikström K, Rurik I, et al. Prevalence and socioeconomic correlates of adult obesity in Europe: the Feel4Diabetes study. Int J Environ Res Public Health. 2022; 19:12572. PMID: 36231871.3. NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in underweight and obesity from 1990 to 2022: a pooled analysis of 3663 population-representative studies with 222 million children, adolescents, and adults. Lancet. 2024; 403:1027–1050. PMID: 38432237.4. Vandevijvere S, De Pauw R, Djojosoeparto S, Gorasso V, Guariguata L, Løvhaug AL, Mialon M, Van Dam I, von Philipsborn P. Upstream determinants of overweight and obesity in Europe. Curr Obes Rep. 2023; 12:417–428. PMID: 37594616.
Article5. Zhu J, Zhang Y, Wu Y, Xiang Y, Tong X, Yu Y, Qiu Y, Cui S, Zhao Q, Wang N, et al. Obesity and dyslipidemia in chinese adults: a cross-sectional study in Shanghai, China. Nutrients. 2022; 14:2321. PMID: 35684121.
Article6. Sun X, Yan AF, Shi Z, Zhao B, Yan N, Li K, Gao L, Xue H, Peng W, Cheskin LJ, et al. Health consequences of obesity and projected future obesity health burden in China. Obesity (Silver Spring). 2022; 30:1724–1751. PMID: 36000246.
Article7. Reid TJ, Korner J. Medical and surgical treatment of obesity. Med Clin North Am. 2022; 106:837–852. PMID: 36154703.
Article8. Müller TD, Blüher M, Tschöp MH, DiMarchi RD. Anti-obesity drug discovery: advances and challenges. Nat Rev Drug Discov. 2022; 21:201–223. PMID: 34815532.9. Perdomo CM, Cohen RV, Sumithran P, Clément K, Frühbeck G. Contemporary medical, device, and surgical therapies for obesity in adults. Lancet. 2023; 401:1116–1130. PMID: 36774932.
Article10. Fink J, Seifert G, Blüher M, Fichtner-Feigl S, Marjanovic G. Obesity surgery. Dtsch Arztebl Int. 2022; 119:70–80. PMID: 34819222.11. Zhu Q, An YA, Scherer PE. Mitochondrial regulation and white adipose tissue homeostasis. Trends Cell Biol. 2022; 32:351–364. PMID: 34810062.
Article12. Li Y, Fromme T. Uncoupling protein 1 does not produce heat without activation. Int J Mol Sci. 2022; 23:2406. PMID: 35269549.
Article13. Crichton PG, Lee Y, Kunji ER. The molecular features of uncoupling protein 1 support a conventional mitochondrial carrier-like mechanism. Biochimie. 2017; 134:35–50. PMID: 28057583.14. Scheele C, Wolfrum C. Brown adipose crosstalk in tissue plasticity and human metabolism. Endocr Rev. 2020; 41:53–65. PMID: 31638161.
Article15. Zhang F, Ma T, Tong X, Liu Y, Cui P, Xu X, Shi J, Hu W, Lu W, Pei Z, et al. Electroacupuncture improves metabolic and ovarian function in a rat model of polycystic ovary syndrome by decreasing white adipose tissue, increasing brown adipose tissue, and modulating the gut microbiota. Acupunct Med. 2022; 40:347–359. PMID: 34892981.
Article16. Cheng L, Wang J, Dai H, Duan Y, An Y, Shi L, Lv Y, Li H, Wang C, Ma Q, et al. Brown and beige adipose tissue: a novel therapeutic strategy for obesity and type 2 diabetes mellitus. Adipocyte. 2021; 10:48–65. PMID: 33403891.
Article17. Saikia R, Joseph J. AMPK: a key regulator of energy stress and calcium-induced autophagy. J Mol Med (Berl). 2021; 99:1539–1551. PMID: 34398293.
Article18. Steinberg GR, Hardie DG. New insights into activation and function of the AMPK. Nat Rev Mol Cell Biol. 2023; 24:255–272. PMID: 36316383.19. Desjardins EM, Steinberg GR. Emerging role of AMPK in brown and beige adipose tissue (BAT): implications for obesity, insulin resistance, and type 2 diabetes. Curr Diab Rep. 2018; 18:80. PMID: 30120579.
Article20. Szewczuk M, Boguszewska K, Kaźmierczak-Barańska J, Karwowski BT. The role of AMPK in metabolism and its influence on DNA damage repair. Mol Biol Rep. 2020; 47:9075–9086. PMID: 33070285.
Article21. Wu YH, Wu YR, Li B, Yan ZY. Cryptotanshinone: a review of its pharmacology activities and molecular mechanisms. Fitoterapia. 2020; 145:104633. PMID: 32445662.
Article22. Wang L, Wang R, Wei GY, Zhang RP, Zhu Y, Wang Z, Wang SM, Du GH. Cryptotanshinone alleviates chemotherapy-induced colitis in mice with colon cancer via regulating fecal-bacteria-related lipid metabolism. Pharmacol Res. 2021; 163:105232. PMID: 33027716.
Article23. Terado T, Kim CJ, Ushio A, Minami K, Tambe Y, Kageyama S, Kawauchi A, Tsunoda T, Shirasawa S, Tanaka H, et al. Cryptotanshinone suppresses tumorigenesis by inhibiting lipogenesis and promoting reactive oxygen species production in KRAS-activated pancreatic cancer cells. Int J Oncol. 2022; 61:108. PMID: 35894141.24. Imran KM, Rahman N, Yoon D, Jeon M, Lee BT, Kim YS. Cryptotanshinone promotes commitment to the brown adipocyte lineage and mitochondrial biogenesis in C3H10T1/2 mesenchymal stem cells via AMPK and p38-MAPK signaling. Biochim Biophys Acta BBAMol Cell Biol Lipids. 2017; 1862:1110–1120.
Article25. Yang W, Han J, Gong S, Zhao J, Yu T, Ma J. Cryptotanshinone suppressed postmenopausal osteoporosis by preventing RANKL-mediated osteoclastogenesis against kidney injury. Evid Based Complement Alternat Med. 2022; 2022:2821984. PMID: 35132325.
Article26. Wang N. Effect of cryptotanshinone on glucose and lipid metabolism and sex hormone levels of rats with polycystic ovary syndrome. Matern Child Health Care China. 2015; 30:3490–3493.27. Ling J, Hua L, Qin Y, Gu T, Jiang S, Zhao J. Perfluorooctane sulfonate promotes hepatic lipid accumulation and steatosis in high-fat diet mice through AMP-activated protein kinase/acetyl-CoA carboxylase (AMPK/ACC) pathway. J Appl Toxicol. 2023; 43:312–322. PMID: 35999056.
Article28. Villarroya F, Cereijo R, Gavaldà-Navarro A, Villarroya J, Giralt M. Inflammation of brown/beige adipose tissues in obesity and metabolic disease. J Intern Med. 2018; 284:492–504. PMID: 29923291.29. Carpi S, Quarta S, Doccini S, Saviano A, Marigliano N, Polini B, Massaro M, Carluccio MA, Calabriso N, Wabitsch M, et al. Tanshinone IIA and Cryptotanshinone counteract inflammation by regulating gene and miRNA expression in human SGBS adipocytes. Biomolecules. 2023; 13:1029. PMID: 37509065.
Article30. Song M, Chen L, Zhang L, Li C, Coffie JW, Fang Z, Zhang L, Wang S, Gao X, Wang H. Cryptotanshinone enhances wound healing in type 2 diabetes with modulatory effects on inflammation, angiogenesis and extracellular matrix remodelling. Pharm Biol. 2020; 58:845–853. PMID: 32870741.
Article31. Tang H, Zeng Q, Tang T, Wei Y, Pu P. Kaempferide improves glycolipid metabolism disorder by activating PPARγ in high-fat-diet-fed mice. Life Sci. 2021; 270:119133. PMID: 33508298.
Article32. Zhang S, Li J, Shi X, Tan X, Si Q. Naringenin activates beige adipocyte browning in high fat diet-fed C57BL/6 mice by shaping the gut microbiota. Food Funct. 2022; 13:9918–9930. PMID: 36053256.
Article33. Wang X, Morris-Natschke SL, Lee KH. New developments in the chemistry and biology of the bioactive constituents of Tanshen. Med Res Rev. 2007; 27:133–148. PMID: 16888751.34. Deng X, Huang SL, Ren J, Pan ZH, Shen Y, Zhou HF, Zuo ZL, Leng Y, Zhao QS. Development and structure-activity relationships of tanshinones as selective 11β-hydroxysteroid dehydrogenase 1 inhibitors. Nat Prod Bioprospect. 2022; 12:36. PMID: 36131216.
Article35. Wang Q, Li H, Tajima K, Verkerke AR, Taxin ZH, Hou Z, Cole JB, Li F, Wong J, Abe I, et al. Post-translational control of beige fat biogenesis by PRDM16 stabilization. Nature. 2022; 609:151–158. PMID: 35978186.
Article36. Chouchani ET, Kazak L, Spiegelman BM. New advances in adaptive thermogenesis: UCP1 and beyond. Cell Metab. 2019; 29:27–37. PMID: 30503034.
Article37. van der Vaart JI, Boon MR, Houtkooper RH. The role of AMPK signaling in brown adipose tissue activation. Cells. 2021; 10:1122. PMID: 34066631.
Article38. Ding X, Jian T, Li J, Lv H, Tong B, Li J, Meng X, Ren B, Chen J. Chicoric acid ameliorates nonalcoholic fatty liver disease via the AMPK/Nrf2/NFκB signaling pathway and restores gut microbiota in high-fat-diet-fed mice. Oxid Med Cell Longev. 2020; 2020:9734560. PMID: 33204402.39. Hu M, Zhang D, Xu H, Zhang Y, Shi H, Huang X, Wang X, Wu Y, Qi Z. Salidroside activates the AMP-activated protein kinase pathway to suppress nonalcoholic steatohepatitis in mice. Hepatology. 2021; 74:3056–3073. PMID: 34292604.
Article40. Entezari M, Hashemi D, Taheriazam A, Zabolian A, Mohammadi S, Fakhri F, Hashemi M, Hushmandi K, Ashrafizadeh M, Zarrabi A, et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: a pre-clinical and clinical investigation. Biomed Pharmacother. 2022; 146:112563. PMID: 35062059.
Article41. Zhang E, Jin L, Wang Y, Tu J, Zheng R, Ding L, Fang Z, Fan M, Al-Abdullah I, Natarajan R, et al. Intestinal AMPK modulation of microbiota mediates crosstalk with brown fat to control thermogenesis. Nat Commun. 2022; 13:1135. PMID: 35241650.
Article42. Diniz TA, de Lima Junior EA, Teixeira AA, Biondo LA, da Rocha LA, Valadão IC, Silveira LS, Cabral-Santos C, de Souza CO, Rosa Neto JC. Aerobic training improves NAFLD markers and insulin resistance through AMPK-PPAR-α signaling in obese mice. Life Sci. 2021; 266:118868. PMID: 33310034.
Article43. Nagappan A, Kim JH, Jung DY, Jung MH. Cryptotanshinone from the Salvia miltiorrhiza bunge attenuates ethanol-induced liver injury by activation of AMPK/SIRT1 and Nrf2 signaling pathways. Int J Mol Sci. 2019; 21:265. PMID: 31906014.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- AMPK and Exercise: Glucose Uptake and Insulin Sensitivity
- Brown Fat and Browning for the Treatment of Obesity and Related Metabolic Disorders
- Quercetin Upregulates Uncoupling Protein 1 in White/Brown Adipose Tissues through Sympathetic Stimulation
- Cryptotanshinone Prevents the Binding of S6K1 to mTOR/Raptor Leading to the Suppression of mTORC1-S6K1 Signaling Activity and Neoplastic Cell Transformation
- Cryptotanshinone but not tanshinone IIA inhibits angiogenesisin vitro
