Vitronectin regulates osteoclastogenesis and bone remodeling in a mouse model of osteoporosis
- Affiliations
-
- 1Graduate School of Humanities and Sciences, Ochanomizu University, Tokyo, Japan
- 2Institute for Human Life Science, Ochanomizu University, Tokyo, Japan
- KMID: 2556575
- DOI: http://doi.org/10.5115/acb.23.251
Abstract
- Vitronectin (VN) is an extracellular matrix protein with a crucial role in regulating bone remodeling. In this study, we aimed to investigate the effect of VN deficiency in a mouse model of osteoporosis induced by ovariectomy (OVX). The findings revealed that the absence of VN led to an increase in the activity of tartrate-resistant acid phosphatase (TRAP), a marker for osteoclasts, in the plasma of OVX-operated mice. TRAP staining further demonstrated that VN deficiency resulted in a higher number of osteoclasts within the femurs of OVX-operated mice. X-ray micro-computed tomography analysis of the femurs in OVX-operated mice indicated that VN deficiency significantly suppressed the OVX-induced increase of marrow area and total volume of bone. Additionally, we assessed structural model index (SMI) and degree of anisotropy (DA) as indices of osteoporosis. The results showed that VN deficiency effectively attenuated the OVX-induced increase in SMI and DA among OVX-operated mice. In summary, our study demonstrates the vital role of VN in regulating osteoclastogenesis and bone remodeling in the mouse model of osteoporosis.
Figure
-
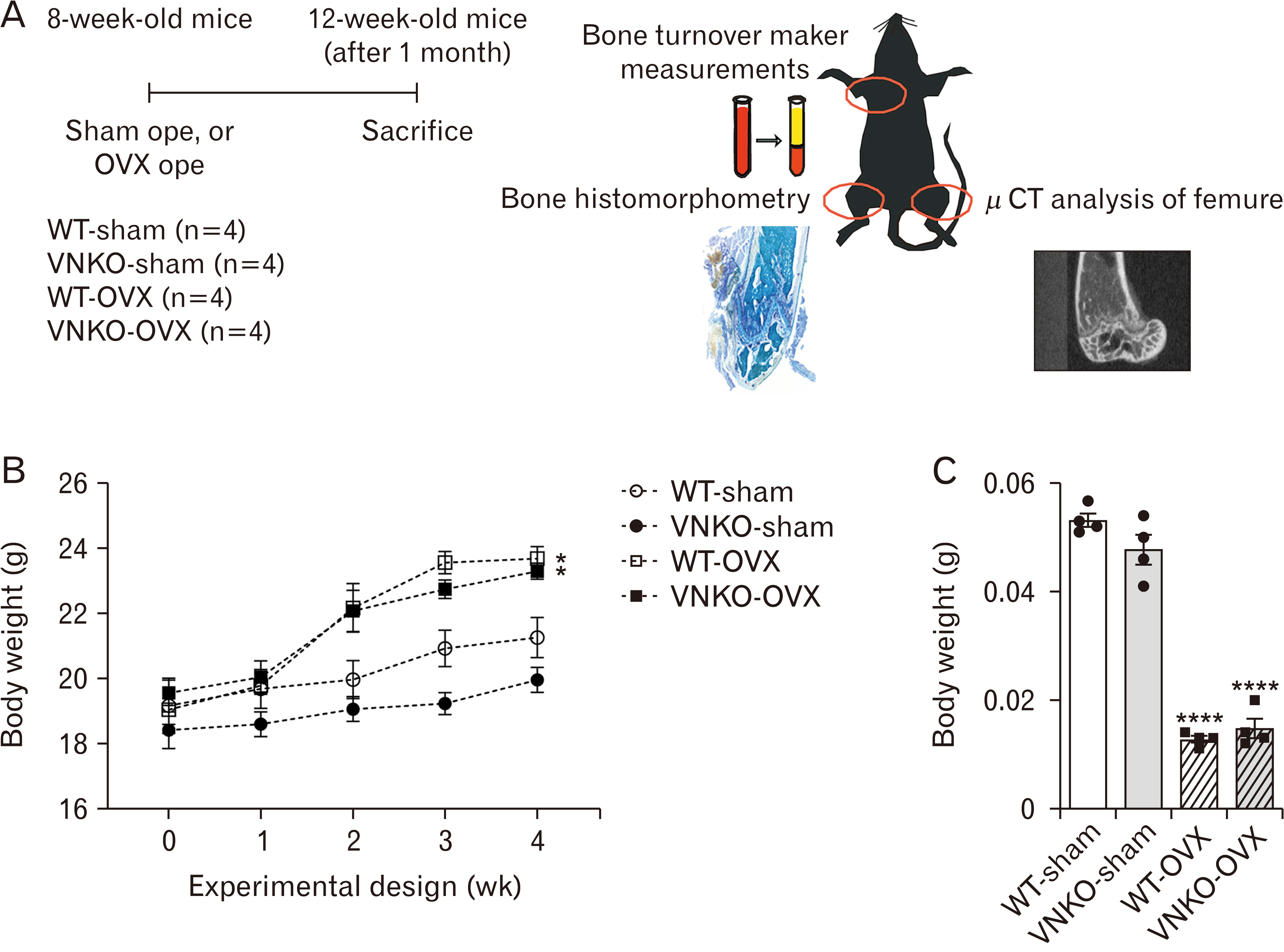
Fig. 1 Effects of ovariectomy on the body and uterine weights of ovariectomized (OVX) mice. (A) Experimental design of this study. Wild-type (WT) female C57BL/6J mice and vitronectin (VN) knockout (KO) female mice were randomly divided into four groups (n=4 mice/group), namely sham-operated WT group (WT-sham), sham-operated VNKO group (VNKO-sham), OVX WT group (WT-OVX), and OVX+VNKO group (VNKO-OVX). The sham and OVX operations were performed in 8-week-old mice, which were sacrificed at 4 weeks after surgery. Plasma was separated from blood by centrifugation and stored at –20°C. The right femur was stored at –20°C for micro-computed tomography (micro-CT) analysis, whereas left femur was fixed in 4% paraformaldehyde at 4°C overnight and subjected to histomorphometry. (B) Body weight change in sham-operated or OVX-operated mice. Body weight was measured in WT-sham, VNKO-sham, WT-OVX, and VNKO-OVX. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. sham-operated group. (C) Uterine weights of sham- or OVX-operated mice. The uterine weight was measured in WT-sham, VNKO-sham, WT-OVX, and VNKO-OVX groups. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: ****P<0.0001 vs. sham-operated group. Dots indicate the results from each animal.

Fig. 2 Localization and expression of vitronectin (VN) in bone remodeling. (A) Immunofluorescent images of femur sections of sham- or ovariectomized (OVX)-operated mice. Top panels: VN (green) and tartrate-resistant acid phosphatase type 5 (TRACP5) (red) expression in the femur sections, with merged images indicating areas of overlap. The arrowheads indicate the sites where VN is close to osteoclasts. Bottom panels: VN (green) and runt related transcription factor 2 (RUNX2) (red) expression in the femur sections. 4’,6-diamidino-2-phenylindole (grey) was used for nuclear counterstaining. Scale bar: 5 μm. (B) Western blot analysis of VN expression in the sera of sham- or OVX-operated mice. Fifty micrograms of protein was loaded into each lane. Relative expression levels of VN protein in wild-type (WT) mice. Data are presented as mean±SEM, n>2. The results were analyzed using the t-test. WT-sham, sham-operated WT group; WT-OVX, OVX WT group; A.U., arbitrary unit; n.s., not significant.
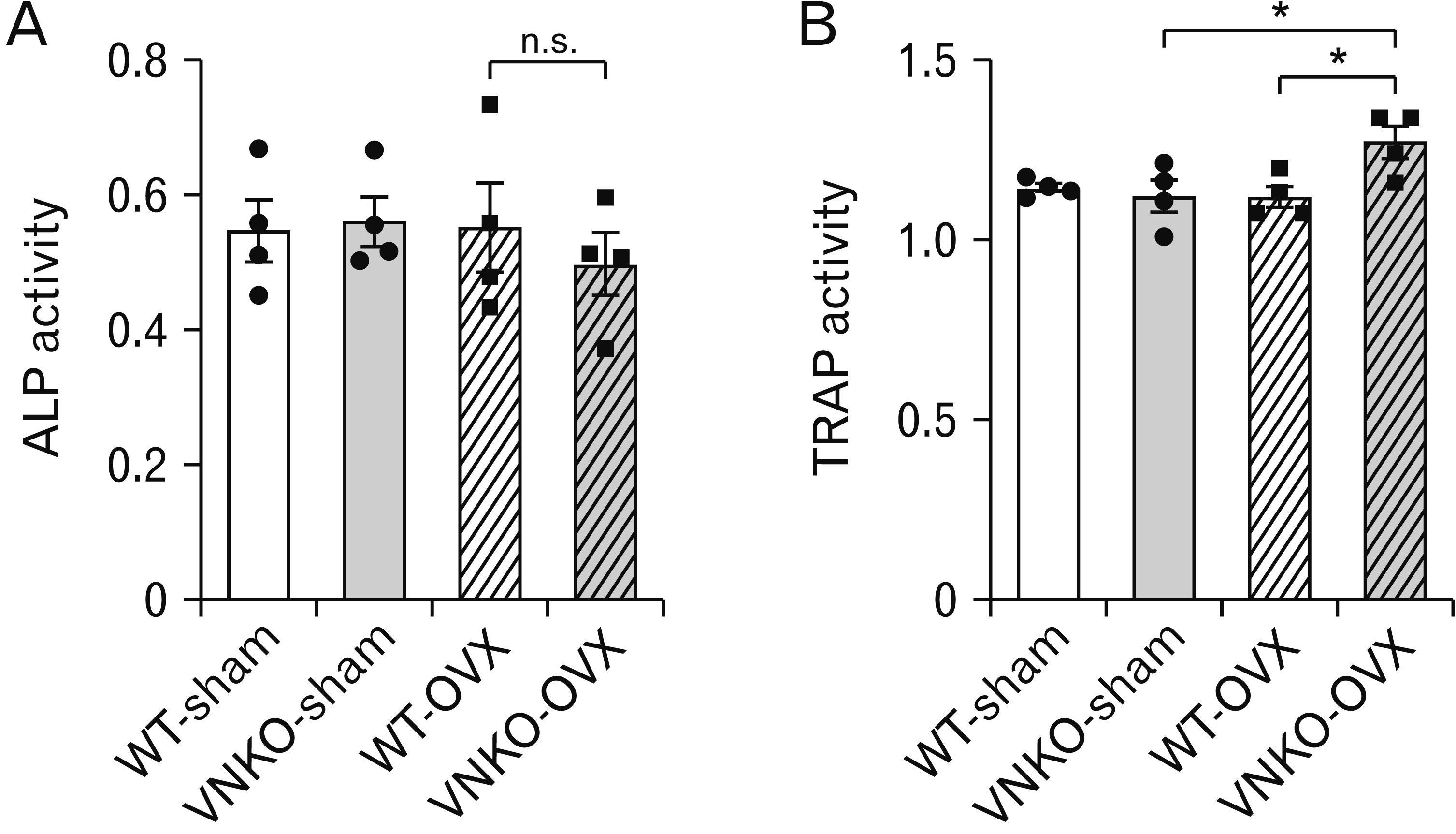
Fig. 3 Effects of vitronectin (VN) deficiency on bone metabolism markers in sham- or ovariectomized (OVX)-operated mice. (A) The activity of plasma alkaline phosphatase (ALP), marker for osteoblasts, in sham- or OVX-operated mice. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA. Dots indicate the results from each animal. (B) The activity of plasma tartrate-resistant acid phosphatase (TRAP), marker for osteoclasts, in sham- or OVX-operated mice. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. OVX+VNKO group (VNKO-OVX). Dots indicate the results from each animal. VNKO, vitronectin knockout; n.s., not significant.

Fig. 4 Alkaline phosphatase (ALP) and tartrate-resistant acid phosphatase (TRAP) staining of femur sections from sham- or ovariectomized (OVX)-operated mice. (A) ALP staining images of the femurs in OVX-operated groups. Scale bar: 100 μm. (B) Bone formation was assessed by measuring the ratio of ALP-positive osteoblast perimeter to bone perimeter (Ob.Pm/B.Pm) in an area 1.0 mm from the growth plate in the distal metaphysis of the femur. Data represent the mean±SEM, n=3. Statistics used one-way ANOVA. Dots indicate the results from each animal. (C) TRAP staining images of the femurs in OVX-operated groups. The arrowheads indicate the activity of osteoclasts. Scale bar: 100 μm. (D) Bone resorption was assessed by measuring the number of TRAP-positive osteoclasts (n>3 nuclei) per bone perimeter (N.Oc/B.Pm) in an area 1.0 mm from the growth plate in the distal metaphysis of the femur. Data represent the mean±SEM, n=3. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. WT-OVX, **P<0.01 vs. WT-sham or VNKO-sham. Dots indicate the results from each animal. WT-sham, sham-operated WT group; VNKO-sham, sham-operated VNKO group; WT-OVX, OVX WT group; VNKO-OVX, OVX+VNKO group; VNKO, vitronectin knockout; n.s., not significant.

Fig. 5 Micro-computed tomography (CT) analysis of femurs in sham- or ovariectomized (OVX)-operated mice. (A) Micro-CT images of the distal femurs obtained after sham- or OVX-operation. Sham-operated wild-type (WT) group (WT-sham), sham-operated VNKO group (VNKO-sham), OVX-operated WT group (WT-OVX), and OVX-operated vitronectin knockout (VNKO) group (VNKO-OVX) were compared; the trabecular bone (orange), the cortical bone (red), and the cortical pore (green). (B, C) Quantitative micro-CT analysis of femurs from sham- or OVX-operated mice. (B) The graph shows parameters of trabecular area of femurs; bone volume (BV), total volume (TV), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp), trabecular number (Tb.N), marrow area (Ma.Ar), degree of anisotropy (DA), and the structural model index (SMI). Data represent the mean±SEM, n>3. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: *P<0.05 vs. WT-OVX, **P<0.01 vs. VNKO-sham, †P<0.05 vs. WT-sham, ††P<0.01 vs. WT-OVX, †††P<0.001 vs. VNKO-sham. Dots indicate the results from each animal. (C) The graph shows parameters of cortical bone area (Ct.Ar) of femurs; Ct.Ar, total cross-sectional area (Tt.Ar), cortical thickness (Ct.Th), and the ratio of Ct.Ar to Tt.Ar. Data represent the mean±SEM, n=4. Statistics used one-way ANOVA and Tukey–Kramer test, P-value: **P<0.01 vs. WT-sham, ***P<0.001 vs. VNKO-sham. Dots indicate the results from each animal.
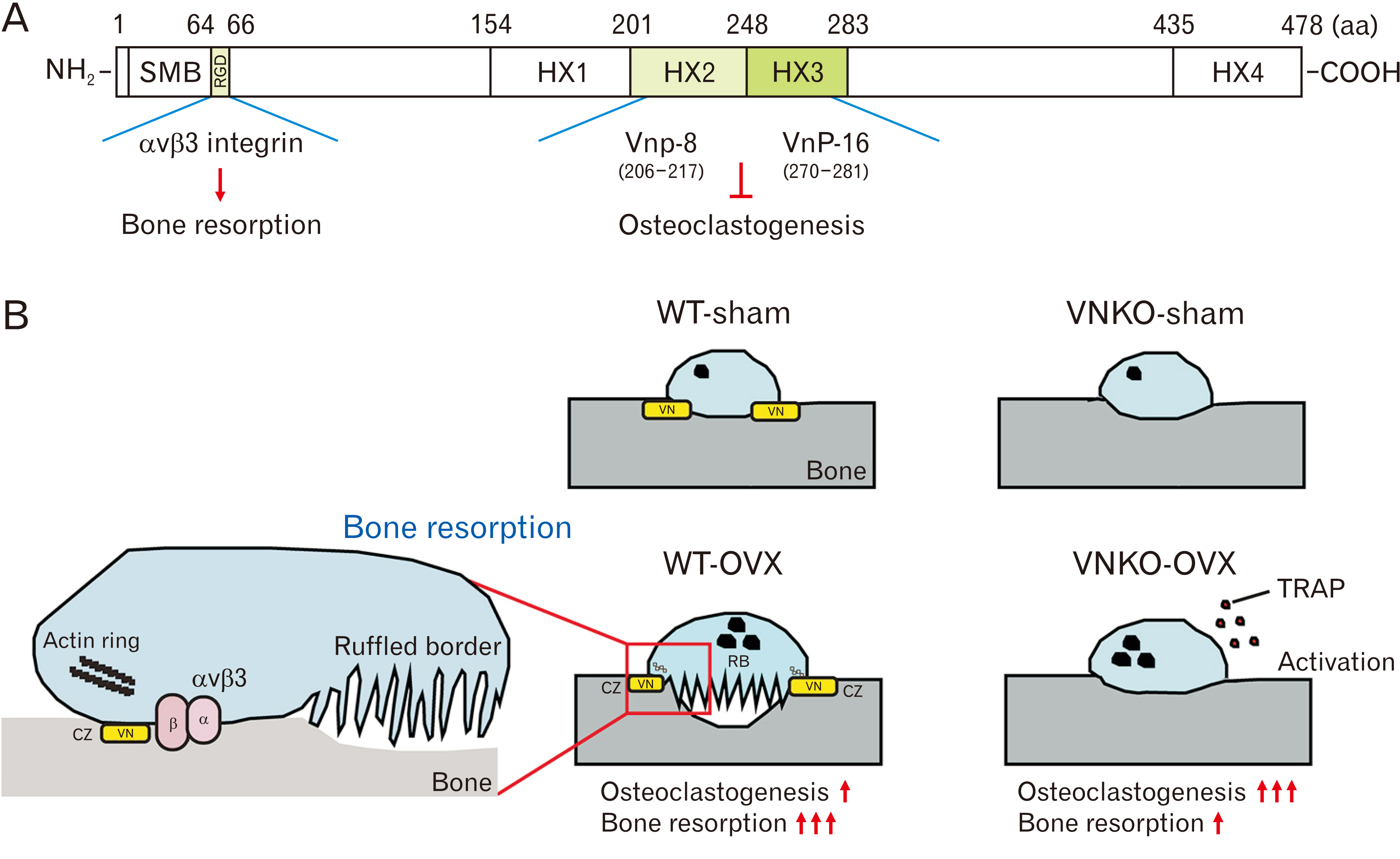
Fig. 6 Functional regions of vitronectin (VN) and the roles in osteoclastogenesis and bone resorption. (A) The amino acid (aa) positions and the domain structure of full-length VN are indicated. The schematic delineates specific regions of VN peptides (VnP-8: residues 206–217 and VnP-16: residues 270–281 in human VN), which are known to inhibit osteoclastogenesis. In contrast, the RGD motif (residues 64–66) is suggested to facilitate the attachment of osteoclasts to VN via αvβ3 integrin on the bone matrix, potentially augmenting bone resorption (B) Diagrams showing the role of VN in osteoclastogenesis and bone resorption in WT-OVX or VNKO-OVX mice. VN regulates osteoclastogenesis and the adhesion of osteoclast to bone surface. In VNKO-OVX mice, VN deficiency increased the activity of tartrate-resistant acid phosphatase (TRAP) and osteoclastogenesis. On the other hand, VN also suppresses osteoclastogenesis. However, VN deficiency suppressed bone resorption in VNKO-OVX mice. VN may regulate the attachment of osteoclasts to bone surface via αvβ3 integrins and induce the actin-rich clear zone (CZ) formation. Furthermore, VN may induce the formation of ruffled borders (RB), leading to bone resorption in WT-OVX mice. WT, wild-type; OVX, ovariectomized; WT-OVX, OVX-operated WT group; VNKO-OVX, OVX-operated vitronectin knockout group; WT-sham, sham-operated WT group; VNKO-sham, sham-operated VNKO group; RGD, Arg-Gly-Asp.
Reference
-
References
1. Wang L, You X, Zhang L, Zhang C, Zou W. 2022; Mechanical regulation of bone remodeling. Bone Res. 10:16. DOI: 10.1038/s41413-022-00190-4. PMID: 35181672. PMCID: PMC8857305.
Article2. Brun P, Scorzeto M, Vassanelli S, Castagliuolo I, Palù G, Ghezzo F, Messina GM, Iucci G, Battaglia V, Sivolella S, Bagno A, Polzonetti G, Marletta G, Dettin M. 2013; Mechanisms underlying the attachment and spreading of human osteoblasts: from transient interactions to focal adhesions on vitronectin-grafted bioactive surfaces. Acta Biomater. 9:6105–15. DOI: 10.1016/j.actbio.2012.12.018. PMID: 23261922.
Article3. Raisz LG. 2005; Pathogenesis of osteoporosis: concepts, conflicts, and prospects. J Clin Invest. 115:3318–25. DOI: 10.1172/JCI27071. PMID: 16322775. PMCID: PMC1297264.
Article4. Tella SH, Gallagher JC. 2014; Prevention and treatment of postmenopausal osteoporosis. J Steroid Biochem Mol Biol. 142:155–70. DOI: 10.1016/j.jsbmb.2013.09.008. PMID: 24176761. PMCID: PMC4187361.
Article5. Klein-Nulend J, van Oers RF, Bakker AD, Bacabac RG. 2015; Bone cell mechanosensitivity, estrogen deficiency, and osteoporosis. J Biomech. 48:855–65. DOI: 10.1016/j.jbiomech.2014.12.007. PMID: 25582356.
Article6. Hayman EG, Pierschbacher MD, Ohgren Y, Ruoslahti E. 1983; Serum spreading factor (vitronectin) is present at the cell surface and in tissues. Proc Natl Acad Sci U S A. 80:4003–7. DOI: 10.1073/pnas.80.13.4003. PMID: 6191326. PMCID: PMC394188.
Article7. Seiffert D. 1996; Detection of vitronectin in mineralized bone matrix. J Histochem Cytochem. 44:275–80. DOI: 10.1177/44.3.8648088. PMID: 8648088.
Article8. Min SK, Kang HK, Jung SY, Jang DH, Min BM. 2018; A vitronectin-derived peptide reverses ovariectomy-induced bone loss via regulation of osteoblast and osteoclast differentiation. Cell Death Differ. 25:268–81. DOI: 10.1038/cdd.2017.153. PMID: 28937683. PMCID: PMC5762842.
Article9. Lee J, Min HK, Park CY, Kang HK, Jung SY, Min BM. 2022; A vitronectin-derived peptide prevents and restores alveolar bone loss by modulating bone re-modelling and expression of RANKL and IL-17A. J Clin Periodontol. 49:799–813. DOI: 10.1111/jcpe.13671. PMID: 35634689.
Article10. Date K, Sakagami H, Yura K. 2021; Regulatory properties of vitronectin and its glycosylation in collagen fibril formation and collagen-degrading enzyme cathepsin K activity. Sci Rep. 11:12023. DOI: 10.1038/s41598-021-91353-6. PMID: 34103584. PMCID: PMC8187593.
Article11. Fuller K, Ross JL, Szewczyk KA, Moss R, Chambers TJ. 2010; Bone is not essential for osteoclast activation. PLoS One. 5:e12837. DOI: 10.1371/journal.pone.0012837. PMID: 20862258. PMCID: PMC2941467.
Article12. Dutra SGV, Felix ACS, Gastaldi AC, De Paula Facioli T, Vieira S, De Souza HCD. 2017; Chronic treatment with angiotensin-converting enzyme inhibitor increases cardiac fibrosis in young rats submitted to early ovarian failure. Auton Neurosci. 206:28–34. DOI: 10.1016/j.autneu.2017.07.001. PMID: 28712539.
Article13. Ikegami H, Kawawa R, Ichi I, Ishikawa T, Koike T, Aoki Y, Fujiwara Y. 2017; Excessive vitamin E intake does not cause bone loss in male or ovariectomized female mice fed normal or high-fat diets. J Nutr. 147:1932–7. DOI: 10.3945/jn.117.248575. PMID: 28835390.
Article14. Li X, Udagawa N, Itoh K, Suda K, Murase Y, Nishihara T, Suda T, Takahashi N. 2002; p38 MAPK-mediated signals are required for inducing osteoclast differentiation but not for osteoclast function. Endocrinology. 143:3105–13. DOI: 10.1210/endo.143.8.8954. PMID: 12130576.
Article15. Feik SA, Thomas CD, Clement JG. 1997; Age-related changes in cortical porosity of the midshaft of the human femur. J Anat. 191(Pt 3):407–16. DOI: 10.1046/j.1469-7580.1997.19130407.x. PMID: 9418997. PMCID: PMC1467697.
Article16. Thomas CD, Feik SA, Clement JG. 2005; Regional variation of intracortical porosity in the midshaft of the human femur: age and sex differences. J Anat. 206:115–25. DOI: 10.1111/j.1469-7580.2005.00384.x. PMID: 15730477. PMCID: PMC1571459.
Article17. Lee SH, Kim JN, Shin KJ, Koh KS, Song WC. 2020; Three-dimensional microstructures of the intracortical canals in the animal model of osteoporosis. Anat Cell Biol. 53:162–8. DOI: 10.5115/acb.19.189. PMID: 32647084. PMCID: PMC7343558.
Article18. Wang B, Dong Y, Tian Z, Chen Y, Dong S. 2021; The role of dendritic cells derived osteoclasts in bone destruction diseases. Genes Dis. 8:401–11. DOI: 10.1016/j.gendis.2020.03.009. PMID: 34179305. PMCID: PMC8209356.
Article19. Väänänen HK, Zhao H, Mulari M, Halleen JM. 2000; The cell biology of osteoclast function. J Cell Sci. 113(Pt 3):377–81. DOI: 10.1242/jcs.113.3.377. PMID: 10639325.
Article20. Takeshita S, Kaji K, Kudo A. 2000; Identification and characterization of the new osteoclast progenitor with macrophage phenotypes being able to differentiate into mature osteoclasts. J Bone Miner Res. 15:1477–88. DOI: 10.1359/jbmr.2000.15.8.1477. PMID: 10934646.
Article21. Tsurukai T, Udagawa N, Matsuzaki K, Takahashi N, Suda T. 2000; Roles of macrophage-colony stimulating factor and osteoclast differentiation factor in osteoclastogenesis. J Bone Miner Metab. 18:177–84. DOI: 10.1007/s007740070018. PMID: 10874596.
Article22. Jung SY, Min BM. 2022; A vitronectin-derived dimeric peptide suppresses osteoclastogenesis by binding to c-Fms and inhibiting M-CSF signaling. Exp Cell Res. 418:113252. DOI: 10.1016/j.yexcr.2022.113252. PMID: 35697077.
Article23. Kang HK, Park CY, Jung SY, Jo SB, Min BM. 2022; A vitronectin-derived peptide restores ovariectomy-induced bone loss by dual regulation of bone remodeling. Tissue Eng Regen Med. 19:1359–76. DOI: 10.1007/s13770-022-00486-w. PMID: 36207661. PMCID: PMC9679078.
Article24. Gramoun A, Azizi N, Sodek J, Heersche JN, Nakchbandi I, Manolson MF. 2010; Fibronectin inhibits osteoclastogenesis while enhancing osteoclast activity via nitric oxide and interleukin-1β-mediated signaling pathways. J Cell Biochem. 111:1020–34. DOI: 10.1002/jcb.22791. PMID: 20672308.
Article25. Geblinger D, Addadi L, Geiger B. 2010; Nano-topography sensing by osteoclasts. J Cell Sci. 123(Pt 9):1503–10. Erratum in: J Cell Sci 2010;123:1814. DOI: 10.1242/jcs.060954. PMID: 20375065. PMCID: PMC2858020.
Article26. Mc Donnell P, Harrison N, Liebschner MA, Mc Hugh PE. 2009; Simulation of vertebral trabecular bone loss using voxel finite element analysis. J Biomech. 42:2789–96. DOI: 10.1016/j.jbiomech.2009.07.038. PMID: 19782987.
Article27. Ding M. 2000; Age variations in the properties of human tibial trabecular bone and cartilage. Acta Orthop Scand Suppl. 292:1–45. DOI: 10.1080/000164700753749791. PMID: 10951715.
Article28. Chambers TJ, Fuller K. 2011; How are osteoclasts induced to resorb bone? Ann N Y Acad Sci. 1240:1–6. DOI: 10.1111/j.1749-6632.2011.06249.x. PMID: 22172032.
Article29. Sanjay A, Houghton A, Neff L, DiDomenico E, Bardelay C, Antoine E, Levy J, Gailit J, Bowtell D, Horne WC, Baron R. 2001; Cbl associates with Pyk2 and Src to regulate Src kinase activity, alpha(v)beta(3) integrin-mediated signaling, cell adhesion, and osteoclast motility. J Cell Biol. 152:181–95. DOI: 10.1083/jcb.152.1.181. PMID: 11149930. PMCID: PMC2193648.30. Lakkakorpi PT, Horton MA, Helfrich MH, Karhukorpi EK, Väänänen HK. 1991; Vitronectin receptor has a role in bone resorption but does not mediate tight sealing zone attachment of osteoclasts to the bone surface. J Cell Biol. 115:1179–86. DOI: 10.1083/jcb.115.4.1179. PMID: 1720122. PMCID: PMC2289948.
Article31. Burgess TL, Qian Y, Kaufman S, Ring BD, Van G, Capparelli C, Kelley M, Hsu H, Boyle WJ, Dunstan CR, Hu S, Lacey DL. 1999; The ligand for osteoprotegerin (OPGL) directly activates mature osteoclasts. J Cell Biol. 145:527–38. DOI: 10.1083/jcb.145.3.527. PMID: 10225954. PMCID: PMC2185088.
Article32. Novack DV, Faccio R. 2011; Osteoclast motility: putting the brakes on bone resorption. Ageing Res Rev. 10:54–61. DOI: 10.1016/j.arr.2009.09.005. PMID: 19788940. PMCID: PMC2888603.
Article33. Stenbeck G. 2002; Formation and function of the ruffled border in osteoclasts. Semin Cell Dev Biol. 13:285–92. DOI: 10.1016/S1084952102000587. PMID: 12243728.
Article34. Davies J, Warwick J, Totty N, Philp R, Helfrich M, Horton M. 1989; The osteoclast functional antigen, implicated in the regulation of bone resorption, is biochemically related to the vitronectin receptor. J Cell Biol. 109(4 Pt 1):1817–26. DOI: 10.1083/jcb.109.4.1817. PMID: 2477382. PMCID: PMC2115816.
Article35. Nesbitt S, Nesbit A, Helfrich M, Horton M. 1993; Biochemical characterization of human osteoclast integrins. Osteoclasts express alpha v beta 3, alpha 2 beta 1, and alpha v beta 1 integrins. J Biol Chem. 268:16737–45. DOI: 10.1016/S0021-9258(19)85479-0. PMID: 8344953.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- A Vitronectin-Derived Peptide Restores Ovariectomy-Induced Bone Loss by Dual Regulation of Bone Remodeling
- Factors and Mechanisms Involved in the Coupling from Bone Resorption to Formation: How Osteoclasts Talk to Osteoblasts
- New Antiresorptive Therapies for Postmenopausal Osteoporosis
- Emerging Anabolic Therapies for Osteoporosis
- High extracellular Ca2+ alone stimulates osteoclast formation but inhibits in the presence of other osteoclastogenic factors
