Docosahexanoic Acid Attenuates Palmitate-Induced Apoptosis by Autophagy Upregulation via GPR120/mTOR Axis in Insulin-Secreting Cells
- Affiliations
-
- 1Institute of Medical Research, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea
- 2Division of Endocrinology and Metabolism, Department of Internal Medicine, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea
- 3Department of Health Sciences and Technology, Samsung Advanced Institute for Health Sciences & Technology, Sungkyunkwan University, Seoul, Korea
- KMID: 2554640
- DOI: http://doi.org/10.3803/EnM.2023.1809
Abstract
- Background
Polyunsaturated fatty acids (PUFAs) reportedly have protective effects on pancreatic β-cells; however, the underlying mechanisms are unknown.
Methods
To investigate the cellular mechanism of PUFA-induced cell protection, mouse insulinoma 6 (MIN6) cells were cultured with palmitic acid (PA) and/or docosahexaenoic acid (DHA), and alterations in cellular signaling and apoptosis were examined.
Results
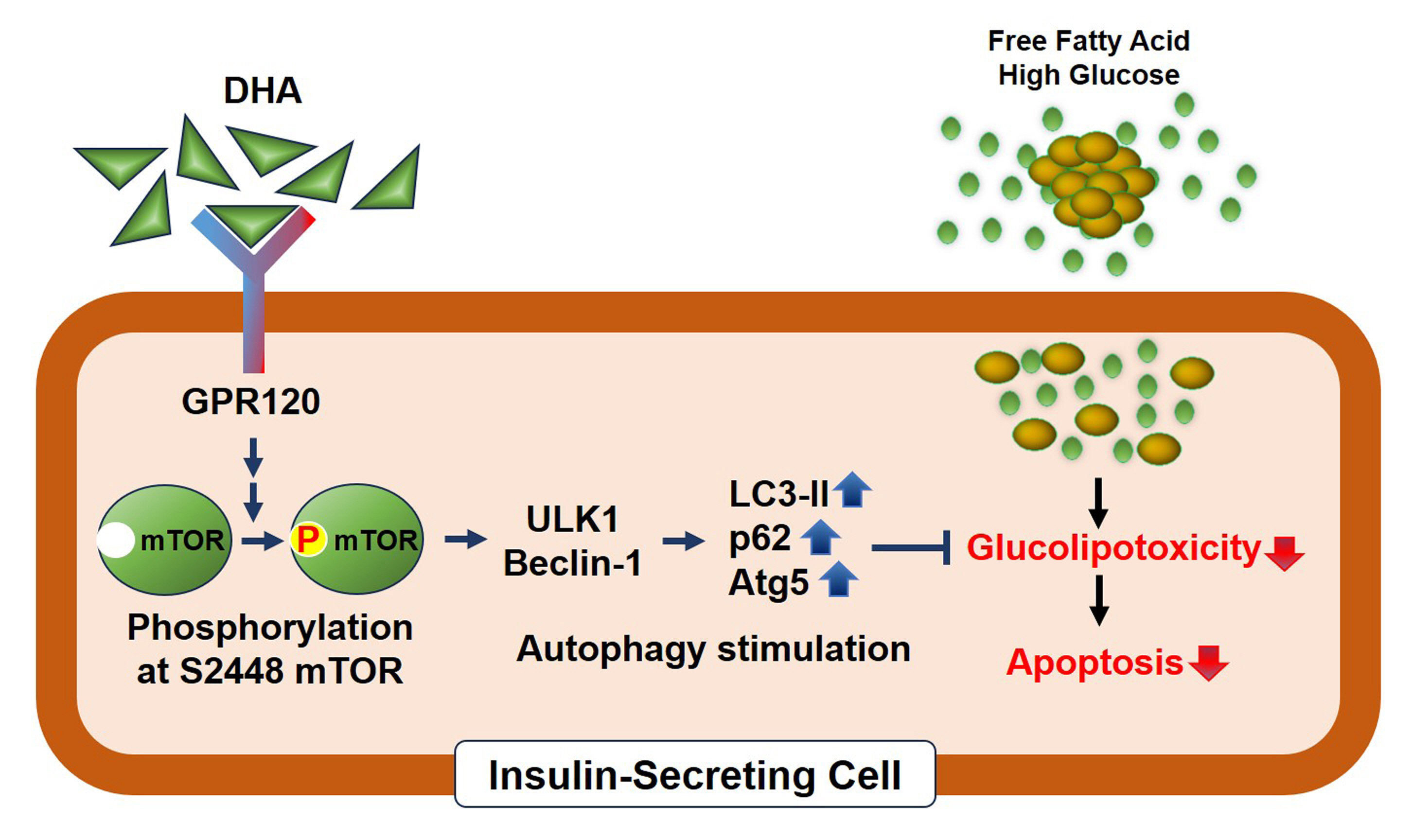
DHA treatment remarkably repressed caspase-3 cleavage and terminal deoxynucleotidyl transferase-mediated UTP nick end labeling (TUNEL)-positive red dot signals in PA-treated MIN6 cells, with upregulation of autophagy, an increase in microtubule- associated protein 1-light chain 3 (LC3)-II, autophagy-related 5 (Atg5), and decreased p62. Upstream factors involved in autophagy regulation (Beclin-1, unc51 like autophagy activating kinase 1 [ULK1], phosphorylated mammalian target of rapamycin [mTOR], and protein kinase B) were also altered by DHA treatment. DHA specifically induced phosphorylation on S2448 in mTOR; however, phosphorylation on S2481 decreased. The role of G protein-coupled receptor 120 (GPR120) in the effect of DHA was demonstrated using a GPR120 agonist and antagonist. Additional treatment with AH7614, a GPR120 antagonist, significantly attenuated DHA-induced autophagy and protection. Taken together, DHA-induced autophagy activation with protection against PA-induced apoptosis mediated by the GPR120/mTOR axis.
Conclusion
These findings indicate that DHA has therapeutic effects on PA-induced pancreatic β-cells, and that the cellular mechanism of β-cell protection by DHA may be a new research target with potential pharmacotherapeutic implications in β-cell protection.
Figure
-
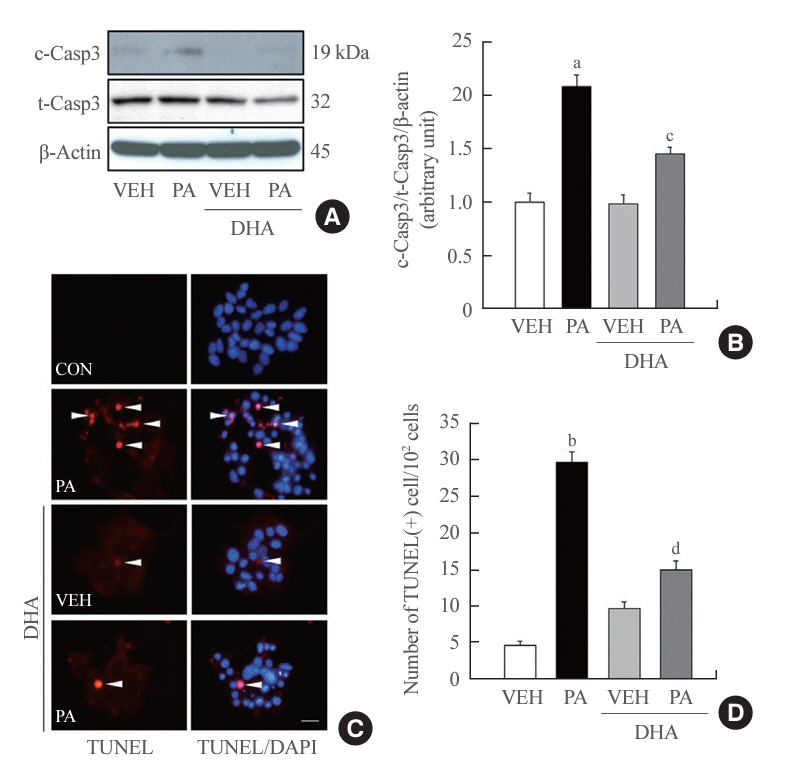
Fig. 1. Docosahexaenoic acid (DHA) prevents palmitic acid (PA)induced apoptosis in insulin-secreting cells. (A, B) Mouse insulinoma 6 (MIN6) cells were incubated with 40 μM DHA in the presence or absence of 500 μM PA for 24 hours. Whole cell lysates were extracted and cleaved caspase-3 (c-Casp3) expression, an apoptosis marker, was measured by immunoblotting analysis. The ratio to total caspase-3 (t-Casp3)/β-actin was described (n=3). (C, D) After treatment with DHA and/or PA, MIN6 cells were fixed for terminal deoxynucleotidyl transferase-mediated UTP nick end labeling (TUNEL) staining assay to observe apoptosis. Apoptotic cells were visualized with red dot signals (white arrows). The nuclei were indicated by 4’,6-diamidino2-phenylindole (DAPI). Scale bars=20 µm. The number of TUNEL-positive cells was counted and described as a ratio to 102 cells (n=3–4). VEH, vehicle; CON, control. aP<0.01, bP<0.001 compared with the VEH group; cP<0.05, dP<0.01 compared with the PA group.
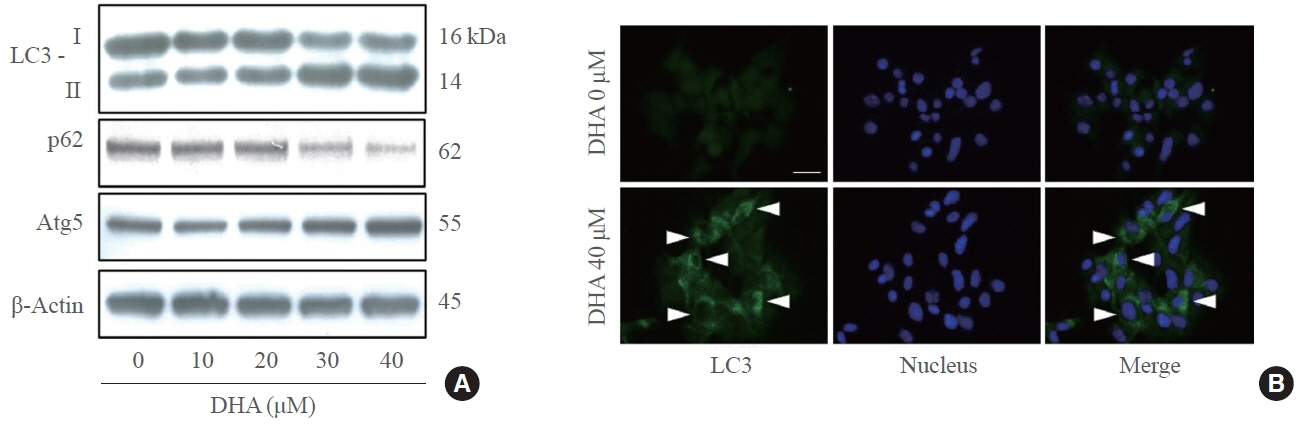
Fig. 2. Docosahexaenoic acid (DHA) upregulates autophagy dose-dependently in insulin-secreting cells. (A) Mouse insulinoma 6 (MIN6) cells exposed to DHA at 0, 10, 20, 30, and 40 μM for 24 hours. Expression of autophagy-related factors including light chain 3 (LC3) I/II, p62, autophagy-related 5 (Atg5) was measured by immunoblotting analysis. (B) MIN6 cells were treated with 40 μM DHA or vehicle for 24 hours, and analyzed by immunofluorescence to morphologically observe the LC3 expression level. LC3 was detected with green fluorescence (white arrowheads), and nuclei were indicated by 4’,6-diamidino2-phenylindole (DAPI). Scale bars=20 µm.
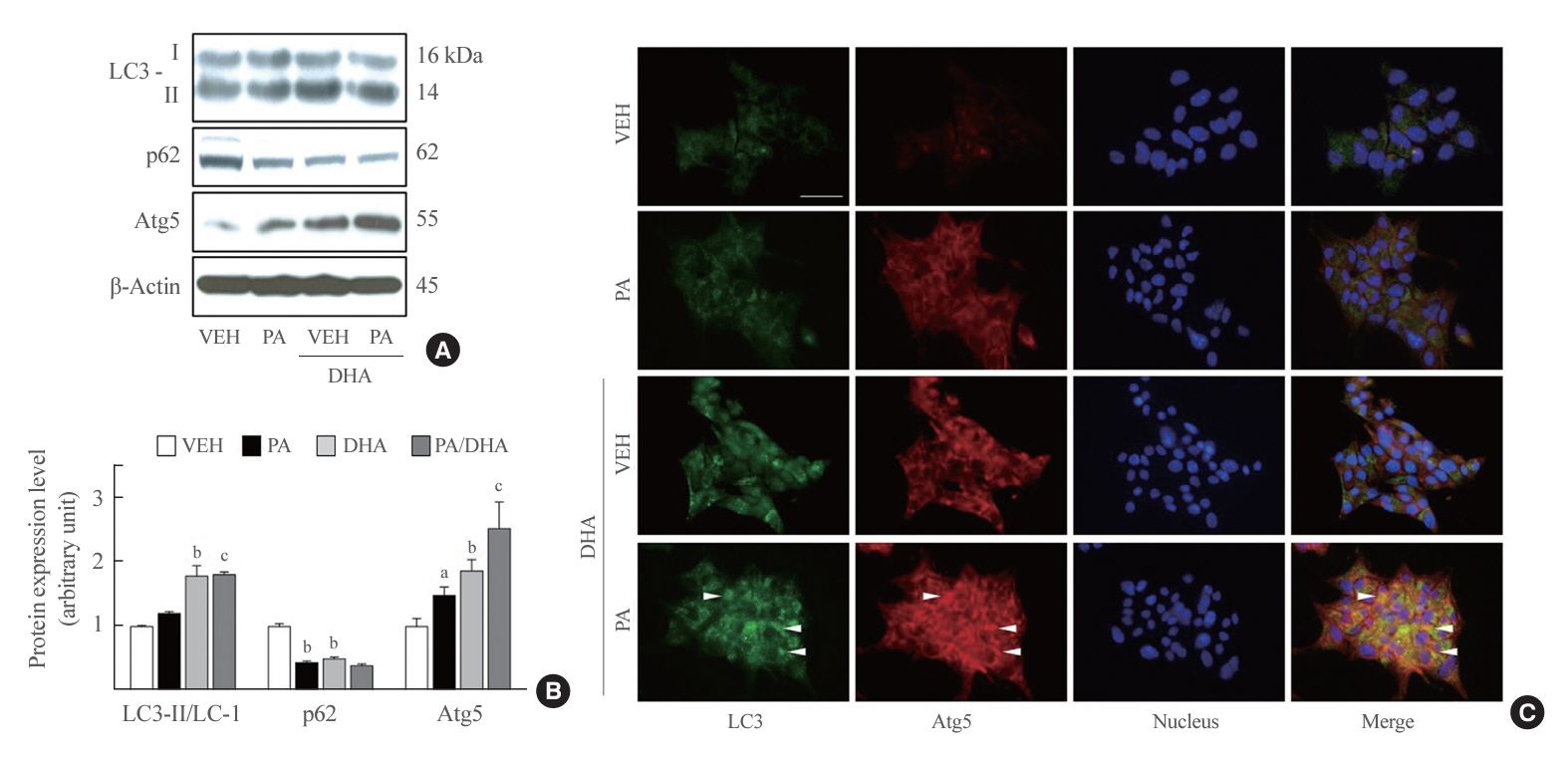
Fig. 3. Docosahexaenoic acid (DHA) upregulates autophagy in a palmitate-treated manner in mouse insulinoma 6 (MIN6) cells. (A, B) Light chain 3 (LC3) I, II, p62, and autophagy-related 5 (Atg5) expression were measured in MIN6 cells after incubation with and without 500 µM palmitate and/or 40 µM DHA for 24 hours. The ratio to β-actin was identified (n=3–4). (C) LC3 and Atg5 expression was detected using double immunofluorescence. LC3 and Atg5 are respectively visualized by green and red fluorescence, and LC3 and Atg5 co-localization is indicated with white arrowheads. Nuclei were marked by 4’,6-diamidino2-phenylindole (DAPI). Scale bar=20 μm. VEH, vehicle; PA, palmitic acid. aP<0.05, bP<0.01 compared with the VEH group; cP<0.01 compared with the PA group.
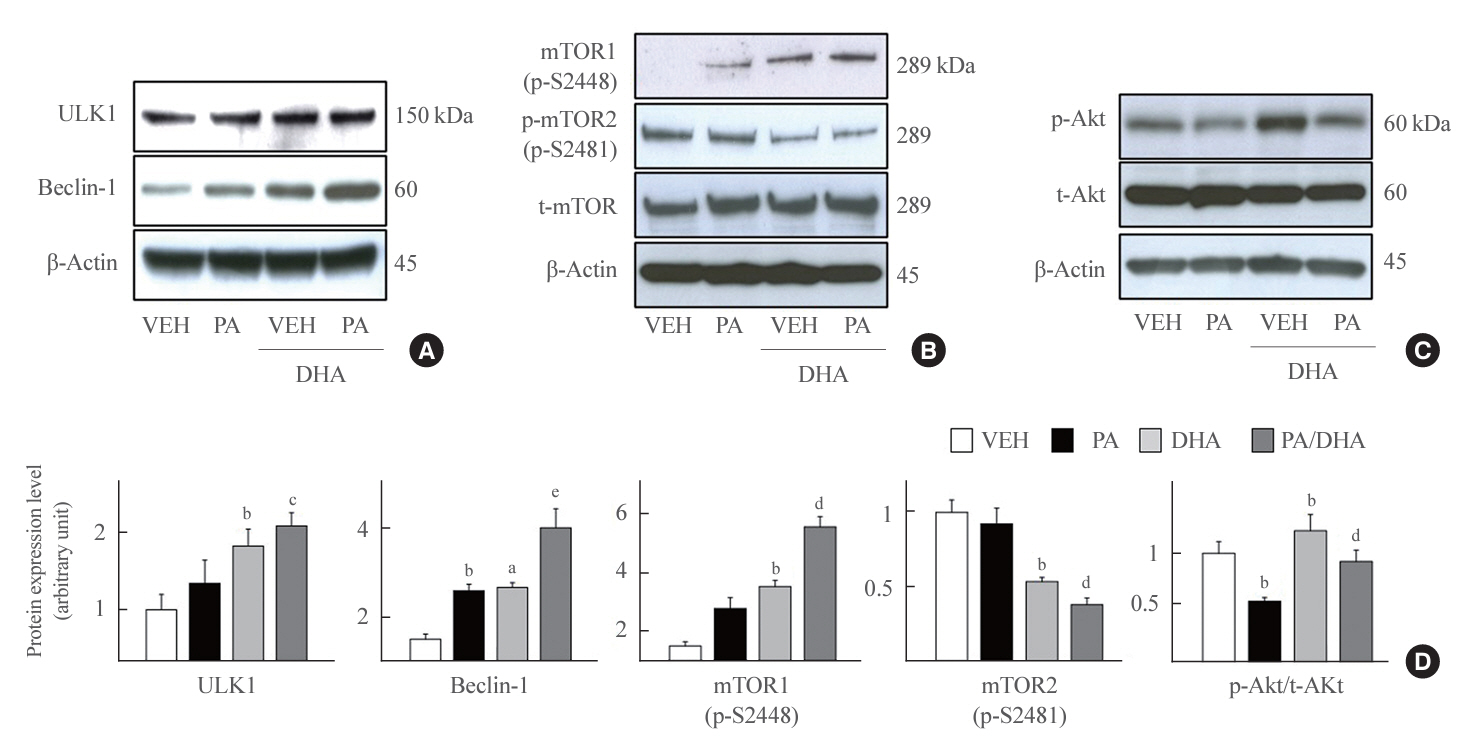
Fig. 4. Docosahexaenoic acid (DHA) affects protein kinase B (Akt)/mammalian target of rapamycin (mTOR)/unc51 like autophagy activating kinase 1 (ULK1) axis. (A, B, C) Mouse insulinoma 6 (MIN6) cells were incubated with 40 μM DHA in the presence or absence of 500 μM palmitate for 24 hours. Whole cell lysates were extracted and analyzed by immunoblotting with antibodies specific for ULK1, Beclin-1, mTOR phosphorylated on S2448 or S2481, or total mTOR (t-mTOR), phosphorylated Akt (p-Akt), total Akt (t-Akt), and β-actin. (D) The ratio to β-actin was described. All values are expressed as the mean±standard error of the mean (n=3–4). VEH, vehicle; PA, palmitic acid. aP<0.05, bP<0.01 compared with the VEH group; cP<0.05, dP<0.01, eP<0.001 compared with the PA group.
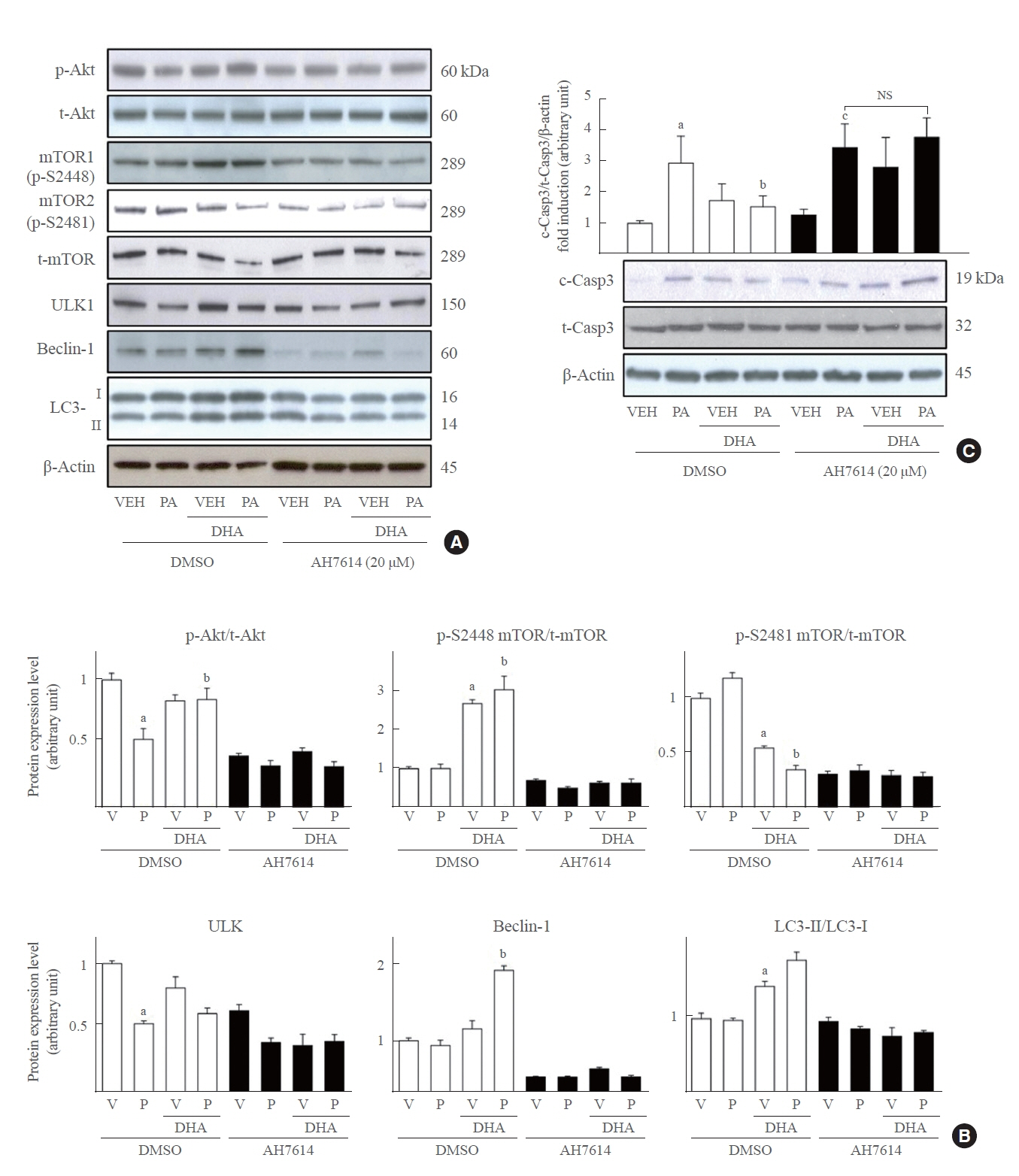
Fig. 5. Inhibition of G protein-coupled receptor 120 (GPR120) strictly blocks the docosahexaenoic acid (DHA) effect in mouse insulinoma 6 (MIN6) cells. MIN6 cells were pretreated with 20 μM AH7614 or dimethyl sulfoxide (DMSO) for 2 hours, and then incubated with palmitate/DHA for 24 hours. (A) Whole cell lysates (WCLs) were extracted and analyzed by immunoblotting with antibodies specific for phosphorylated protein kinase B (p-Akt), total Akt (t-Akt), mammalian target of rapamycin (mTOR) phosphorylated on S2448 or S2481, or total mTOR (t-mTOR), unc51 like autophagy activating kinase 1 (ULK1), Beclin-1 light chain 3 (LC3) II/I, and β-actin. (B) The ratio of each factor’s band intensity to β-actin was described. (C) Cleaved caspase-3 (c-Casp3) and total caspase-3 (t-Casp3) were measured by immunoblotting analysis, and the level of c-Casp3 was normalized to t-Casp3/β-actin. All values are expressed as the mean±standard error of the mean (n=3). VEH (V), vehicle; PA (P), palmitic acid; NS, non-significant. aP<0.01 compared with the VEH group of DMSO set; bP<0.05 compared with the PA group of DMSO set; cP<0.01 compared with VEH group of AH7614 set.
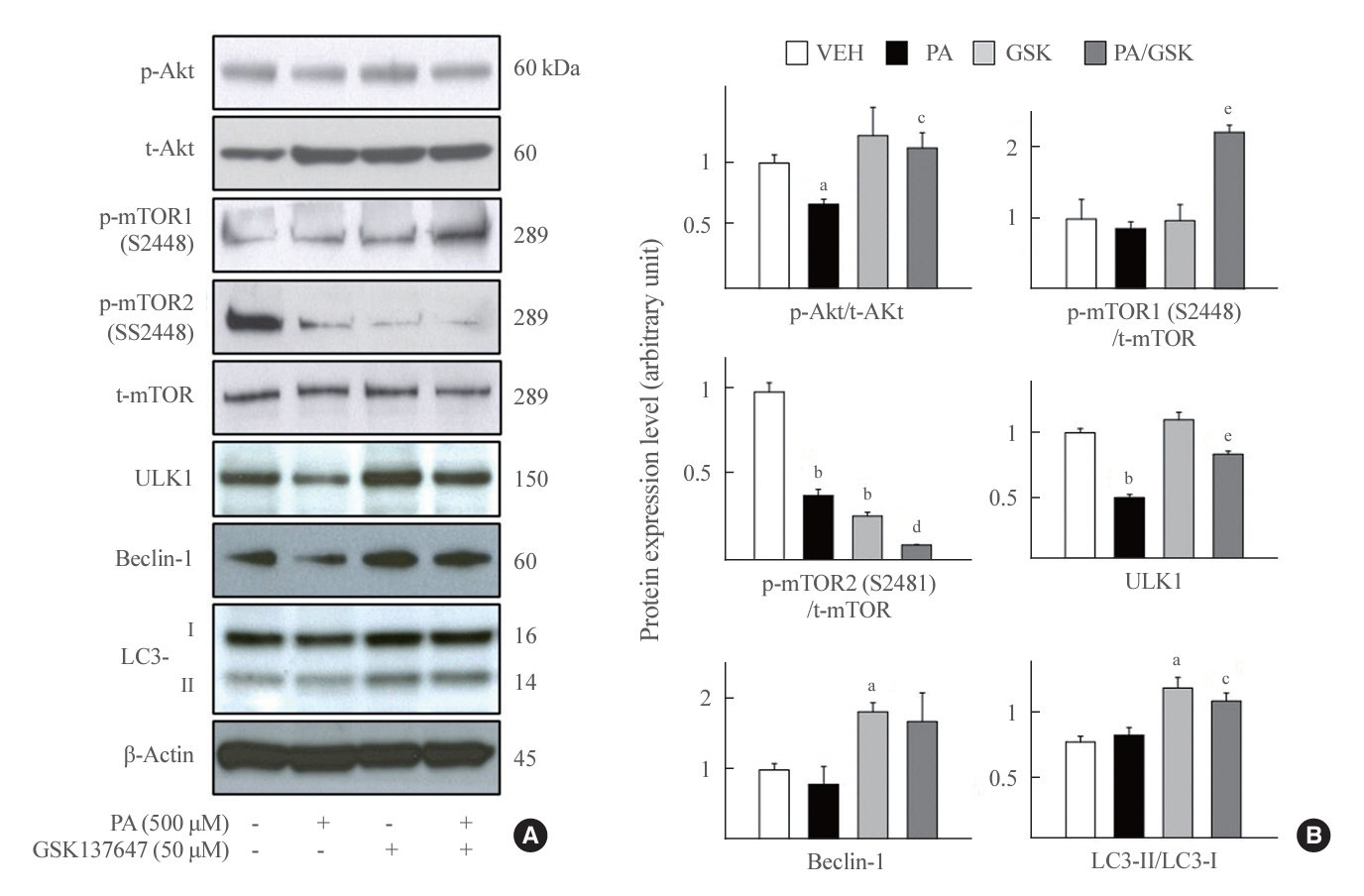
Fig. 6. Autophagy activation by G protein-coupled receptor 120 (GPR120) agonist via protein kinase B (Akt)/mammalian target of rapamycin (mTOR) axis. Mouse insulinoma 6 (MIN6) cells were incubated with or without 500 μM palmitate and/or 50 μM GSK137647 for 24 hours. (A, B) Whole cell lysates were extracted and analyzed by immunoblotting with antibodies specific for phosphorylated Akt (p-Akt), total Akt (t-Akt), mTOR phosphorylated on S2448 or S2481, or total mTOR (t-mTOR), unc51 like autophagy activating kinase 1 (ULK1), Beclin-1 light chain 3 (LC3) I/II, and β-actin. All values are expressed as the mean±standard error of the mean (n=3–4). VEH, vehicle; PA, palmitic acid. aP<0.05, bP<0.001 compared with the VEH group; cP<0.05, dP<0.01, eP<0.001 compared with the PA group.
Reference
-
1. Martin de Santa Olalla L, Sanchez Muniz FJ, Vaquero MP. N-3 fatty acids in glucose metabolism and insulin sensitivity. Nutr Hosp. 2009; 24:113–27.2. Fedor D, Kelley DS. Prevention of insulin resistance by n-3 polyunsaturated fatty acids. Curr Opin Clin Nutr Metab Care. 2009; 12:138–46.
Article3. Djousse L, Biggs ML, Lemaitre RN, King IB, Song X, Ix JH, et al. Plasma omega-3 fatty acids and incident diabetes in older adults. Am J Clin Nutr. 2011; 94:527–33.
Article4. Rhee EJ. The influence of obesity and metabolic health on vascular health. Endocrinol Metab (Seoul). 2022; 37:1–8.
Article5. Shimabukuro M, Higa M, Zhou YT, Wang MY, Newgard CB, Unger RH. Lipoapoptosis in beta-cells of obese prediabetic fa/fa rats. Role of serine palmitoyltransferase overexpression. J Biol Chem. 1998; 273:32487–90.6. El-Assaad W, Buteau J, Peyot ML, Nolan C, Roduit R, Hardy S, et al. Saturated fatty acids synergize with elevated glucose to cause pancreatic beta-cell death. Endocrinology. 2003; 144:4154–63.7. Maedler K, Spinas GA, Lehmann R, Sergeev P, Weber M, Fontana A, et al. Glucose induces beta-cell apoptosis via upregulation of the Fas receptor in human islets. Diabetes. 2001; 50:1683–90.8. Shimabukuro M, Zhou YT, Levi M, Unger RH. Fatty acidinduced beta cell apoptosis: a link between obesity and diabetes. Proc Natl Acad Sci U S A. 1998; 95:2498–502.9. Lucena CF, Roma LP, Graciano MF, Veras K, Simoes D, Curi R, et al. Omega-3 supplementation improves pancreatic islet redox status: in vivo and in vitro studies. Pancreas. 2015; 44:287–95.10. Bellini L, Campana M, Rouch C, Chacinska M, Bugliani M, Meneyrol K, et al. Protective role of the ELOVL2/docosahexaenoic acid axis in glucolipotoxicity-induced apoptosis in rodent beta cells and human islets. Diabetologia. 2018; 61:1780–93.
Article11. Kebede MA, Alquier T, Latour MG, Poitout V. Lipid receptors and islet function: therapeutic implications? Diabetes Obes Metab. 2009; 11 Suppl 4:10–20.
Article12. Davenport AP, Alexander SP, Sharman JL, Pawson AJ, Benson HE, Monaghan AE, et al. International Union of Basic and Clinical Pharmacology. LXXXVIII. G protein-coupled receptor list: recommendations for new pairings with cognate ligands. Pharmacol Rev. 2013; 65:967–86.
Article13. Mobraten K, Haug TM, Kleiveland CR, Lea T. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco2 cells. Lipids Health Dis. 2013; 12:101.
Article14. Kang S, Huang J, Lee BK, Jung YS, Im E, Koh JM, et al. Omega-3 polyunsaturated fatty acids protect human hepatoma cells from developing steatosis through FFA4 (GPR120). Biochim Biophys Acta Mol Cell Biol Lipids. 2018; 1863:105–16.
Article15. Ren Z, Chen L, Wang Y, Wei X, Zeng S, Zheng Y, et al. Activation of the omega-3 fatty acid receptor GPR120 protects against focal cerebral ischemic injury by preventing inflammation and apoptosis in mice. J Immunol. 2019; 202:747–59.
Article16. Oh DY, Talukdar S, Bae EJ, Imamura T, Morinaga H, Fan W, et al. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell. 2010; 142:687–98.
Article17. Wang Y, Xie T, Zhang D, Leung PS. GPR120 protects lipotoxicity-induced pancreatic β-cell dysfunction through regulation of PDX1 expression and inhibition of islet inflammation. Clin Sci (Lond). 2019; 133:101–16.
Article18. Glick D, Barth S, Macleod KF. Autophagy: cellular and molecular mechanisms. J Pathol. 2010; 221:3–12.
Article19. Mizushima N. Autophagy: process and function. Genes Dev. 2007; 21:2861–73.
Article20. Gonzalez CD, Lee MS, Marchetti P, Pietropaolo M, Towns R, Vaccaro MI, et al. The emerging role of autophagy in the pathophysiology of diabetes mellitus. Autophagy. 2011; 7:2–11.
Article21. Hong SW, Lee J, Kim MJ, Moon SJ, Kwon H, Park SE, et al. clusterin protects lipotoxicity-induced apoptosis via upregulation of autophagy in insulin-secreting cells. Endocrinol Metab (Seoul). 2020; 35:943–53.
Article22. Quan W, Hur KY, Lim Y, Oh SH, Lee JC, Kim KH, et al. Autophagy deficiency in beta cells leads to compromised unfolded protein response and progression from obesity to diabetes in mice. Diabetologia. 2012; 55:392–403.
Article23. Gehrmann W, Elsner M, Lenzen S. Role of metabolically generated reactive oxygen species for lipotoxicity in pancreatic β-cells. Diabetes Obes Metab. 2010; 12 Suppl 2:149–58.24. Kharroubi I, Ladriere L, Cardozo AK, Dogusan Z, Cnop M, Eizirik DL. Free fatty acids and cytokines induce pancreatic beta-cell apoptosis by different mechanisms: role of nuclear factor-kappaB and endoplasmic reticulum stress. Endocrinology. 2004; 145:5087–96.25. El-Assaad W, Joly E, Barbeau A, Sladek R, Buteau J, Maestre I, et al. Glucolipotoxicity alters lipid partitioning and causes mitochondrial dysfunction, cholesterol, and ceramide deposition and reactive oxygen species production in INS832/13 sscells. Endocrinology. 2010; 151:3061–73.26. Zhou C, Wu Z, Du W, Que H, Wang Y, Ouyang Q, et al. Recycling of autophagosomal components from autolysosomes by the recycler complex. Nat Cell Biol. 2022; 24:497–512.
Article27. Schlafli AM, Adams O, Galvan JA, Gugger M, Savic S, Bubendorf L, et al. Prognostic value of the autophagy markers LC3 and p62/SQSTM1 in early-stage non-small cell lung cancer. Oncotarget. 2016; 7:39544–55.
Article28. Otomo C, Metlagel Z, Takaesu G, Otomo T. Structure of the human ATG12~ATG5 conjugate required for LC3 lipidation in autophagy. Nat Struct Mol Biol. 2013; 20:59–66.
Article29. Li XD, He SS, Wan TT, Li YB. Liraglutide protects palmitate-induced INS-1 cell injury by enhancing autophagy mediated via FoxO1. Mol Med Rep. 2021; 23:147.
Article30. Chu KY, O’Reilly L, Mellet N, Meikle PJ, Bartley C, Biden TJ. Oleate disrupts cAMP signaling, contributing to potent stimulation of pancreatic β-cell autophagy. J Biol Chem. 2019; 294:1218–29.
Article31. Heras-Sandoval D, Perez-Rojas JM, Hernandez-Damian J, Pedraza-Chaverri J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal. 2014; 26:2694–701.
Article32. Alers S, Loffler AS, Wesselborg S, Stork B. Role of AMPKmTOR-Ulk1/2 in the regulation of autophagy: cross talk, shortcuts, and feedbacks. Mol Cell Biol. 2012; 32:2–11.
Article33. Russell RC, Tian Y, Yuan H, Park HW, Chang YY, Kim J, et al. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat Cell Biol. 2013; 15:741–50.
Article34. Biden TJ, Boslem E, Chu KY, Sue N. Lipotoxic endoplasmic reticulum stress, β cell failure, and type 2 diabetes mellitus. Trends Endocrinol Metab. 2014; 25:389–98.
Article35. Bhaswant M, Poudyal H, Brown L. Mechanisms of enhanced insulin secretion and sensitivity with n-3 unsaturated fatty acids. J Nutr Biochem. 2015; 26:571–84.
Article36. Baynes HW, Mideksa S, Ambachew S. The role of polyunsaturated fatty acids (n-3 PUFAs) on the pancreatic β-cells and insulin action. Adipocyte. 2018; 7:81–7.
Article37. Liu W, Zheng Q, Zhu M, Liu X, Liu J, Lu Y, et al. Regulatory effects of N-3 PUFAs on pancreatic β-cells and insulinsensitive tissues. Curr Drug Metab. 2021; 22:1017–34.
Article38. Chen Y, Xu C, Yan T, Yu C, Li Y. ω-3 Fatty acids reverse lipotoxity through induction of autophagy in nonalcoholic fatty liver disease. Nutrition. 2015; 31:1423–9.
Article39. Liu W, Xu L, Wang X, Zhang D, Sun G, Wang M, et al. PRDX1 activates autophagy via the PTEN-AKT signaling pathway to protect against cisplatin-induced spiral ganglion neuron damage. Autophagy. 2021; 17:4159–81.
Article40. Riahi Y, Wikstrom JD, Bachar-Wikstrom E, Polin N, Zucker H, Lee MS, et al. Autophagy is a major regulator of beta cell insulin homeostasis. Diabetologia. 2016; 59:1480–91.
Article41. Muralidharan C, Conteh AM, Marasco MR, Crowder JJ, Kuipers J, de Boer P, et al. Pancreatic beta cell autophagy is impaired in type 1 diabetes. Diabetologia. 2021; 64:865–77.
Article42. Wang Y, Zhang H. Regulation of autophagy by mTOR signaling pathway. Adv Exp Med Biol. 2019; 1206:67–83.
Article43. Copp J, Manning G, Hunter T. TORC-specific phosphorylation of mammalian target of rapamycin (mTOR): phosphoSer2481 is a marker for intact mTOR signaling complex 2. Cancer Res. 2009; 69:1821–7.
Article44. Soliman GA, Acosta-Jaquez HA, Dunlop EA, Ekim B, Maj NE, Tee AR, et al. mTOR Ser-2481 autophosphorylation monitors mTORC-specific catalytic activity and clarifies rapamycin mechanism of action. J Biol Chem. 2010; 285:7866–79.
Article45. Rosner M, Siegel N, Valli A, Fuchs C, Hengstschlager M. mTOR phosphorylated at S2448 binds to raptor and rictor. Amino Acids. 2010; 38:223–8.
Article46. Tripathy S, Jump DB. Elovl5 regulates the mTORC2-Akt-FOXO1 pathway by controlling hepatic cis-vaccenic acid synthesis in diet-induced obese mice. J Lipid Res. 2013; 54:71–84.
Article47. Liu R, Chen L, Wang Z, Zheng X, Hou Z, Zhao D, et al. Omega-3 polyunsaturated fatty acids prevent obesity by improving tricarboxylic acid cycle homeostasis. J Nutr Biochem. 2021; 88:108503.
Article48. Zhang D, So WY, Wang Y, Wu SY, Cheng Q, Leung PS. Insulinotropic effects of GPR120 agonists are altered in obese diabetic and obese non-diabetic states. Clin Sci (Lond). 2017; 131:247–60.
Article49. Chen J, Wang D, Zong Y, Yang X. DHA protects hepatocytes from oxidative injury through GPR120/ERK-mediated mitophagy. Int J Mol Sci. 2021; 22:5675.
Article50. Gao B, Han YH, Wang L, Lin YJ, Sun Z, Lu WG, et al. Eicosapentaenoic acid attenuates dexamethasome-induced apoptosis by inducing adaptive autophagy via GPR120 in murine bone marrow-derived mesenchymal stem cells. Cell Death Dis. 2016; 7:e2235.
Article51. Williams-Bey Y, Boularan C, Vural A, Huang NN, Hwang IY, Shan-Shi C, et al. Omega-3 free fatty acids suppress macrophage inflammasome activation by inhibiting NF-κB activation and enhancing autophagy. PLoS One. 2014; 9:e97957.
Article52. Moon S, Jung HS. Endoplasmic reticulum stress and dysregulated autophagy in human pancreatic beta cells. Diabetes Metab J. 2022; 46:533–42.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Clusterin Protects Lipotoxicity-Induced Apoptosis via Upregulation of Autophagy in Insulin-Secreting Cells
- Decreased Expression and Induced Nucleocytoplasmic Translocation of Pancreatic and Duodenal Homeobox 1 in INS-1 Cells Exposed to High Glucose and Palmitate
- Regulatory Role of Autophagy in Globular Adiponectin-Induced Apoptosis in Cancer Cells
- Autophagy in Diabetes
- Dasatinib induces apoptosis and autophagy by suppressing the PI3K/Akt/mTOR pathway in bladder cancer cells
