Quercetin inhibits body weight gain and adipogenesis via matrix metalloproteinases in mice fed a highfat diet
- Affiliations
-
- 1Department of Food Science and Nutrition, Dankook University, Chungnam 31116, Korea
- KMID: 2542841
- DOI: http://doi.org/10.4162/nrp.2023.17.3.438
Abstract
- BACKGROUND/OBJECTIVES
Limited studies reported that quercetin inhibited adipogenesis and neovascularization by inhibiting matrix metalloproteinases (MMPs) activity, but such mechanisms have not been elucidated in animal experiments. In this study, we investigated the inhibitory effects of quercetin on weight gain and adipose tissue growth through the regulation of mRNA expressions of adipogenic transcription factors and MMPs in mice fed a high-fat diet (HFD).
MATERIALS/METHODS
Five-wk-old C57BL/6J mice were fed a normal diet (ND), HFD, HFD containing 0.05% of quercetin (HFQ0.05), or HFD containing 0.15% of quercetin (HFQ0.15) for 16 wks. Glycerol-3-phosphate dehydrogenase (GPDH) activity was measured using a commercial kit. The mRNA expressions of transcription factors related to adipocyte differentiation were determined by real-time polymerase chain reaction (PCR). The mRNA expressions of MMPs and concentrations of MMPs were measured by real-time PCR and enzyme-linked immunosorbent assay kit, respectively.
RESULTS
Quercetin intake reduced body weight gain and epididymal adipose tissue weights (P < 0.05). GPDH activity was higher in the HFD group than in the ND group but lower in the quercetin groups (P < 0.05). The mRNA expressions of CCAAT/enhancer binding protein β (C/EBPβ), C/EBPα, peroxisome proliferator-activated receptor γ, and fatty acid-binding protein 4 were lower in the quercetin groups than in the HFD group (P < 0.05). Similarly, the mRNA expression and concentrations of MMP-2 and MMP-9 were significantly lower in the quercetin groups than in the HFD group (P < 0.05).
CONCLUSION
The study confirms that quercetin suppresses body weight gain and adipogenesis by inhibiting transcription factors related to adipocyte differentiation and MMPs (MMP-2 and MMP-9), in mice fed a HFD.
Keyword
Figure
-
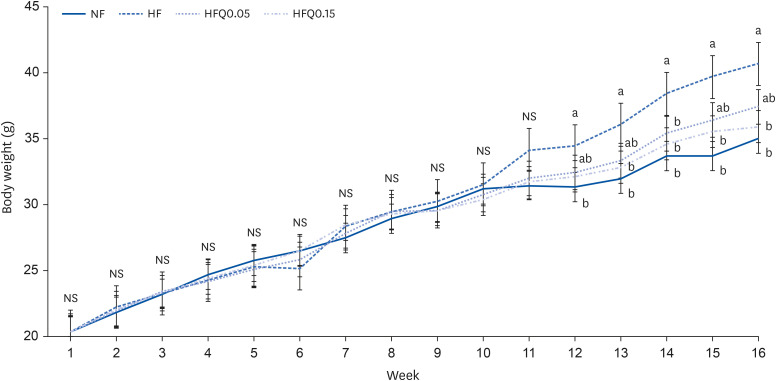
Fig. 1 Body weight changes in the experimental groups.Lines represent mean weights ± SEs of the experimental groups. Five-wk-old C57BL/6J mice (Daehan Biolink) were randomly allocated to 4 experimental groups: normal fat diet (ND); high fat diet (HFD); high fat diet + quercetin 0.05% (HFQ0.05); and high fat diet + quercetin 0.15% (HFQ0.15).NS, no significant intergroup difference.Different symbols above lines for each wk indicate significant intergroup differences at P < 0.05 as determined by Duncans multiple range test.
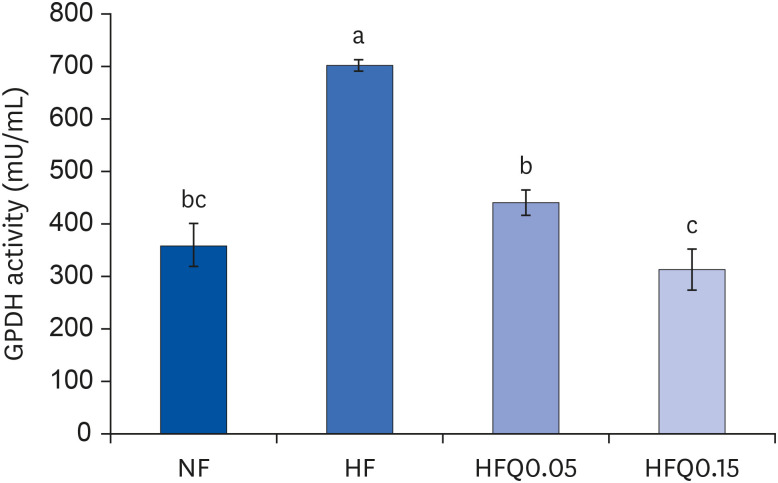
Fig. 2 Effect of 16 weeks of quercetin administration on GPDH activity in adipose tissues.Bars represent means ± SEs of the experimental groups. Five-wk-old C57BL/6J mice (Daehan Biolink) were randomly allocated to 4 experimental groups:normal fat diet (ND); high fat diet (HFD); high fat diet + quercetin 0.05% (HFQ0.05); and high fat diet + quercetin 0.15% (HFQ0.15).GPDH, glycerol-3-phosphate dehydrogenase.Different letters above bars indicate significant intergroup differences at P < 0.05 as determined by Duncans multiple range test.
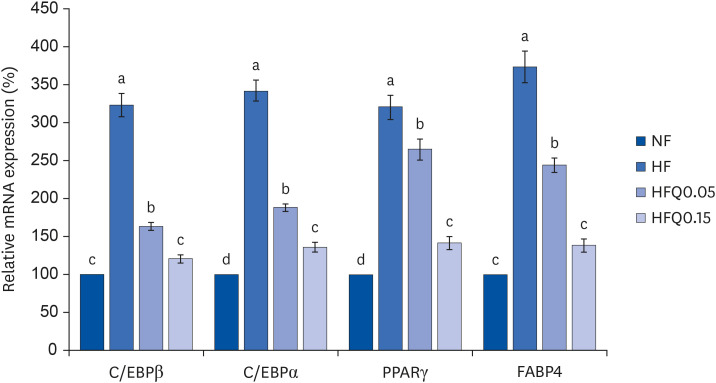
Fig. 3 Effects of 16 weeks of quercetin administration on the mRNA expressions of transcription factors related to adipocyte differentiation in the liver.Total RNA was isolated using TRI-Reagent, and cDNA was synthesized using SuperScript II reverse transcriptase from total RNA. Real-time PCR with SYBR Green was performed using standard procedures to assess the mRNA expressions in liver samples obtained from each group. β-Actin levels were used to ensure equal loadings. Applied Biosystems StepOne software v2.1 was used. Bars represent means ± SEs of the experimental groups. Five-wk-old C57BL/6J mice (Daehan Biolink) were randomly allocated to 4 experimental groups: normal fat diet (ND); high fat diet (HFD); high fat diet + quercetin 0.05% (HFQ0.05); and high fat diet + quercetin 0.15% (HFQ0.15).PCR, polymerase chain reaction; C/EBPβ, CCAT/enhancer-binding protein β; C/EBPα, CCAT/enhancer-binding protein α; PPARγ, peroxisome proliferator activated receptor γ; FABP4, fatty acid binding protein 4.Different letters above bars indicate significant intergroup differences (P < 0.05 as determined by Duncans multiple range test).
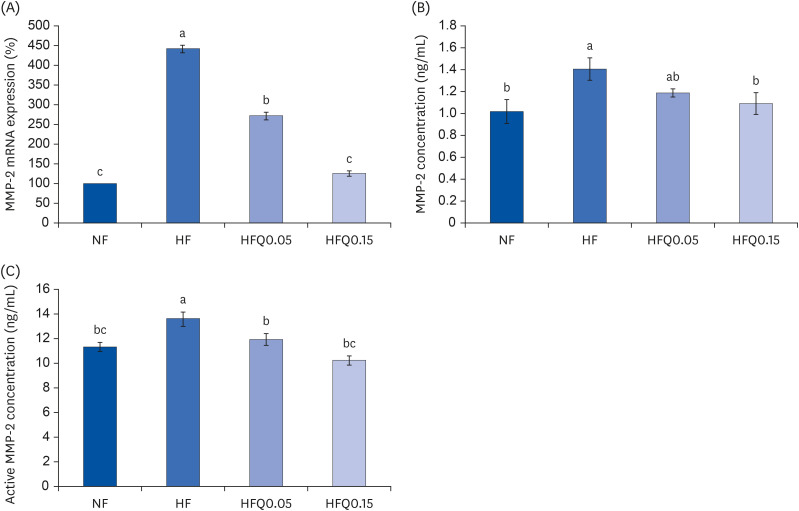
Fig. 4 Effects of 16 weeks of quercetin administration on mRNA expression, concentrations of MMP-2 in adipose tissues.To measure the mRNA expression of MMP-2, total RNA was isolated using TRI-Reagent, and cDNA was synthesized using total RNA and SuperScript II reverse transcriptase. Real-time PCR with SYBR Green was performed using standard procedures to assess mRNA expressions in epididymal fat pad samples. (A) β-Actin levels were used to ensure equal loadings. Applied Biosystems StepOne software v2.1 was used. (B) MMP-2 protein levels were measured using the MMP-2 ELISA kit (MBS722437, MyBioSouce. (C) MMP-2 activity was measured using the MMP-2 Biotrack activity assay system kit (RPN 2631; GE Healthcare. Bars represent means ± SEs of the experimental groups. Five-wk-old C57BL/6J mice (Daehan Biolink) were randomly allocated to 4 experimental groups: normal fat diet (ND); high fat diet (HFD); high fat diet + quercetin 0.05% (HFQ0.05); and high fat diet + quercetin 0.15% (HFQ0.15).MMP, matrix metalloproteinase.Different letters above bars indicate significant intergroup differences (P < 0.05 as determined by Duncans multiple range test).
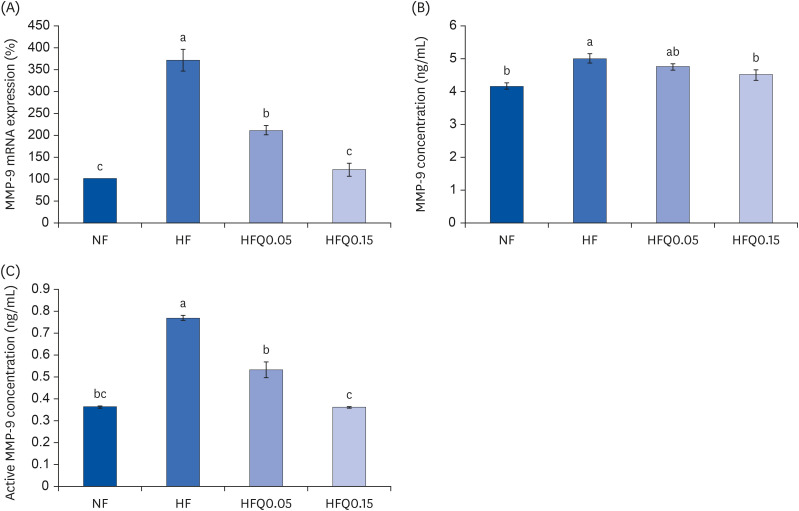
Fig. 5 Effects of 16 weeks of quercetin administration on the mRNA expression, concentrations of MMP-9 in adipose tissues.To measure the mRNA expression of MMP-9, total RNA was isolated using TRI-Reagent, and cDNA was synthesized using total RNA with SuperScript II reverse transcriptase. Real-time PCR with SYBR Green was performed using standard procedures to determine mRNA expressions in epididymal fat pad samples. (A) β-Actin levels were used to ensure equal loadings. Applied Biosystems StepOne software v2.1 was used. (B) MMP-9 levels were measured using the MMP-9 ELISA kit (MBS720876, MyBioSource, and (C) MMP-9 activities were measured using an MMP-9 activity assay kit (QuickZyme BioScience). Bars represent means ± SEs of the experimental groups. Five-wk-old C57BL/6J mice (Daehan Biolink) were randomly allocated to 4 experimental groups: normal fat diet (ND); high fat diet (HFD); high fat diet + quercetin 0.05% (HFQ0.05); and high fat diet + quercetin 0.15% (HFQ0.15).MMP, matrix metalloproteinase; PCR, polymerase chain reaction; ELISA, enzyme-linked immunosorbent assay.Different letters above bars indicate significant intergroup differences at P < 0.05 as determined by Duncans multiple range tests.
Reference
-
1. Korea Centers for Disease Control and Prevention. Trends in prevalence of obesity, 2011–2020. Public Health Wkly Rep. 2022; 15:2693–2694.2. Marcelin G, Silveira AL, Martins LB, Ferreira AV, Clément K. Deciphering the cellular interplays underlying obesity-induced adipose tissue fibrosis. J Clin Invest. 2019; 129:4032–4040. PMID: 31498150.3. Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis. Nature. 2011; 473:298–307. PMID: 21593862.4. Guo L, Li X, Tang QQ. Transcriptional regulation of adipocyte differentiation: a central role for CCAAT/enhancer-binding protein (C/EBP) β. J Biol Chem. 2015; 290:755–761. PMID: 25451943.5. Ali AT, Hochfeld WE, Myburgh R, Pepper MS. Adipocyte and adipogenesis. Eur J Cell Biol. 2013; 92:229–236. PMID: 23876739.6. Bellafiore M, Battaglia G, Bianco A, Farina F, Palma A, Paoli A. The involvement of MMP-2 and MMP-9 in heart exercise-related angiogenesis. J Transl Med. 2013; 11:283. PMID: 24195673.7. Kruegel J, Miosge N. Basement membrane components are key players in specialized extracellular matrices. Cell Mol Life Sci. 2010; 67:2879–2895. PMID: 20428923.8. Shin SS, Yoon M. Regulation of obesity by antiangiogenic herbal medicines. Molecules. 2020; 25:4549. PMID: 33020443.9. Balaji M, Ganjayi MS, Hanuma Kumar GE, Parim BN, Mopuri R, Dasari S. A review on possible therapeutic targets to contain obesity: the role of phytochemicals. Obes Res Clin Pract. 2016; 10:363–380. PMID: 26740473.10. Hsu CL, Yen GC. Induction of cell apoptosis in 3T3-L1 pre-adipocytes by flavonoids is associated with their antioxidant activity. Mol Nutr Food Res. 2006; 50:1072–1079. PMID: 17039455.11. Hsu CL, Yen GC. Phenolic compounds: evidence for inhibitory effects against obesity and their underlying molecular signaling mechanisms. Mol Nutr Food Res. 2008; 52:53–61. PMID: 18081207.12. Ku HC, Chang HH, Liu HC, Hsiao CH, Lee MJ, Hu YJ, Hung PF, Liu CW, Kao YH. Green tea (-)-epigallocatechin gallate inhibits insulin stimulation of 3T3-L1 preadipocyte mitogenesis via the 67-kDa laminin receptor pathway. Am J Physiol Cell Physiol. 2009; 297:C121–C132. PMID: 19176763.13. Ambati S, Yang JY, Rayalam S, Park HJ, Della-Fera MA, Baile CA. Ajoene exerts potent effects in 3T3-L1 adipocytes by inhibiting adipogenesis and inducing apoptosis. Phytother Res. 2009; 23:513–518. PMID: 19051208.14. Boccellino M, D’Angelo S. Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int J Mol Sci. 2020; 21:5642. PMID: 32781724.15. Yang JY, Della-Fera MA, Rayalam S, Ambati S, Hartzell DL, Park HJ, Baile CA. Enhanced inhibition of adipogenesis and induction of apoptosis in 3T3-L1 adipocytes with combinations of resveratrol and quercetin. Life Sci. 2008; 82:1032–1039. PMID: 18433793.16. Ahn J, Lee H, Kim S, Park J, Ha T. The anti-obesity effect of quercetin is mediated by the AMPK and MAPK signaling pathways. Biochem Biophys Res Commun. 2008; 373:545–549. PMID: 18586010.17. Hong SY, Ha AW, Kim W. Effects of quercetin on cell differentiation and adipogenesis in 3T3-L1 adipocytes. Nutr Res Pract. 2021; 15:444–455. PMID: 34349878.18. Song SM, Jung SM, Choi SJ, Kim WK. Quercetin inhibits adipocyte differentiation via matrix metalloproteinases in 3T3-L1 Preadipocytes. J Korean Soc Food Sci and Nutr. 2022; 51:311–332.19. Nijhawans P, Behl T, Bhardwaj S. Angiogenesis in obesity. Biomed Pharmacother. 2020; 126:110103. PMID: 32200253.20. Park BS, Han SH, Lee JY, Chung YS. Evaluation of in vivo genotoxicity of plant flavonoids, quercetin and isoquercetin. J Food Hyg Saf. 2016; 31:356–364.21. Moon J, Do HJ, Kim OY, Shin MJ. Antiobesity effects of quercetin-rich onion peel extract on the differentiation of 3T3-L1 preadipocytes and the adipogenesis in high fat-fed rats. Food Chem Toxicol. 2013; 58:347–354. PMID: 23684756.22. Porras D, Nistal E, Martínez-Flórez S, Pisonero-Vaquero S, Olcoz JL, Jover R, González-Gallego J, García-Mediavilla MV, Sánchez-Campos S. Protective effect of quercetin on high-fat diet-induced non-alcoholic fatty liver disease in mice is mediated by modulating intestinal microbiota imbalance and related gut-liver axis activation. Free Radic Biol Med. 2017; 102:188–202. PMID: 27890642.23. Jia Q, Cao H, Shen D, Li S, Yan L, Chen C, Xing S, Dou F. Quercetin protects against atherosclerosis by regulating the expression of PCSK9, CD36, PPARγ, LXRα and ABCA1. Int J Mol Med. 2019; 44:893–902. PMID: 31524223.24. Lee JS, Cha YJ, Lee KH, Yim JE. Onion peel extract reduces the percentage of body fat in overweight and obese subjects: a 12-week, randomized, double-blind, placebo-controlled study. Nutr Res Pract. 2016; 10:175–181. PMID: 27087901.25. Jung CH, Cho I, Ahn J, Jeon TI, Ha TY. Quercetin reduces high-fat diet-induced fat accumulation in the liver by regulating lipid metabolism genes. Phytother Res. 2013; 27:139–143. PMID: 22447684.26. Bae CR, Park YK, Cha YS. Quercetin-rich onion peel extract suppresses adipogenesis by down-regulating adipogenic transcription factors and gene expression in 3T3-L1 adipocytes. J Sci Food Agric. 2014; 94:2655–2660. PMID: 24634340.27. Zhao X, Hu H, Wang C, Bai L, Wang Y, Wang W, Wang J. A comparison of methods for effective differentiation of the frozen-thawed 3T3-L1 cells. Anal Biochem. 2019; 568:57–64. PMID: 30594506.28. Boughanem H, Cabrera-Mulero A, Millán-Gómez M, Garrido-Sánchez L, Cardona F, Tinahones FJ, Moreno-Santos I, Macías-González M. Transcriptional analysis of FOXO1, C/EBP-α and PPAR-γ2 genes and their association with obesity-related insulin resistance. Genes (Basel). 2019; 10:706. PMID: 31547433.29. Moseti D, Regassa A, Kim WK. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int J Mol Sci. 2016; 17:124. PMID: 26797605.30. Cristancho AG, Lazar MA. Forming functional fat: a growing understanding of adipocyte differentiation. Nat Rev Mol Cell Biol. 2011; 12:722–734. PMID: 21952300.31. Zhang R, Qin X, Zhang T, Li Q, Zhang J, Zhao J. Astragalus polysaccharide improves insulin sensitivity via AMPK activation in 3T3-L1 adipocytes. Molecules. 2018; 23:2711. PMID: 30347867.32. Bouloumié A, Sengenès C, Portolan G, Galitzky J, Lafontan M. Adipocyte produces matrix metalloproteinases 2 and 9: involvement in adipose differentiation. Diabetes. 2001; 50:2080–2086. PMID: 11522674.33. Bauters D, Scroyen I, Van Hul M, Lijnen HR. Gelatinase A (MMP-2) promotes murine adipogenesis. Biochim Biophys Acta. 2015; 1850:1449–1456. PMID: 25869489.34. Bosco DB, Roycik MD, Jin Y, Schwartz MA, Lively TJ, Zorio DA, Sang QA. A new synthetic matrix metalloproteinase inhibitor reduces human mesenchymal stem cell adipogenesis. PLoS One. 2017; 12:e0172925. PMID: 28234995.35. Park BY, Lee H, Woo S, Yoon M, Kim J, Hong Y, Lee HS, Park EK, Hahm JC, Kim JW, et al. Reduction of adipose tissue mass by the angiogenesis inhibitor ALS-L1023 from melissa officinalis. PLoS One. 2015; 10:e0141612. PMID: 26599360.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Inhibitory Effect of Quercetin on Adipose Tissue Inflammation in Mice Fed on a High-fat Diet
- Capsanthin Inhibits both Adipogenesis in 3T3-L1 Preadipocytes and Weight Gain in High-Fat Diet-Induced Obese Mice
- Arctiin inhibits adipogenesis in 3T3-L1 cells and decreases adiposity and body weight in mice fed a high-fat diet
- Effects of quercetin on the improvement of lipid metabolism through regulating hepatic AMPK and microRNA-21 in high cholesterol diet-fed mice
- Effects of quercetin nanoemulsion on SIRT1 activation and mitochondrial biogenesis in the skeletal muscle of high-fat diet-fed mice
