β1,4-Galactosyltransferase V Modulates Breast Cancer Stem Cells through Wnt/β-catenin Signaling Pathway
- Affiliations
-
- 1Key Laboratory of Marine Drugs, Chinese Ministry of Education, School of Medicine and Pharmacy, Ocean University of China, Qingdao, China
- 2Laboratory for Marine Drugs and Bioproducts of Qingdao National Laboratory for Marine Science and Technology, Qingdao, China
- 3Open Studio for Druggability Research of Marine Natural Products, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, China
- KMID: 2507935
- DOI: http://doi.org/10.4143/crt.2020.093
Abstract
- Purpose
Breast cancer stem cells (BCSCs) contribute to the initiation, development, and recurrence of breast carcinomas. β1,4-Galactosyltransferase V (B4GalT5), which catalyzes the addition of galactose to GlcNAcβ1-4Man of N-glycans, is involved in embryogenesis. However, its role in the modulation of BCSCs remains unknown.
Materials and Methods
The relationship between B4GalT5 and breast cancer stemness was investigated by online clinical databases and immunohistochemistry analysis. Mammosphere formation, fluorescence-activated cell sorting (FACS), and in-vivo assays were used to evaluate B4GalT5 expression in BCSCs and its effect on BCSCs. B4GalT5 regulation of Wnt/β-catenin signaling was examined by immunofluorescence and Ricinus communis agglutinin I pull-down assays. Cell surface biotinylation and FACS assays were performed to assess the association of cell surface B4GalT5 and BCSCs.
Results
B4GalT5, but not other B4GalTs, was highly correlated with BCSC markers and poor prognosis. B4GalT5 significantly increased the stem cell marker aldehyde dehydrogenase 1A1 (ALDH1A1) and promoted the production of CD44+CD24–/low cells and the formation of mammospheres. Furthermore, B4GalT5 overexpression resulted in dramatic tumor growth in vivo. Mechanistically, B4GalT5 modified and protected Frizzled-1 from degradation via the lysosomal pathway, promoting Wnt/β-catenin signaling which was hyperactivated in BCSCs. B4GalT5, located on the surface of a small subset of breast carcinoma cells, was not responsible for the stemness of BCSCs.
Conclusion
B4GalT5 modulates the stemness of breast cancer through glycosylation modification to stabilize Frizzled-1 and activate Wnt/β-catenin signaling independent of its cell surface location. Our studies highlight a previously unknown role of B4GalT5 in regulating the stemness of breast cancer and provide a potential drug target for anticancer drug development.
Keyword
Figure
-
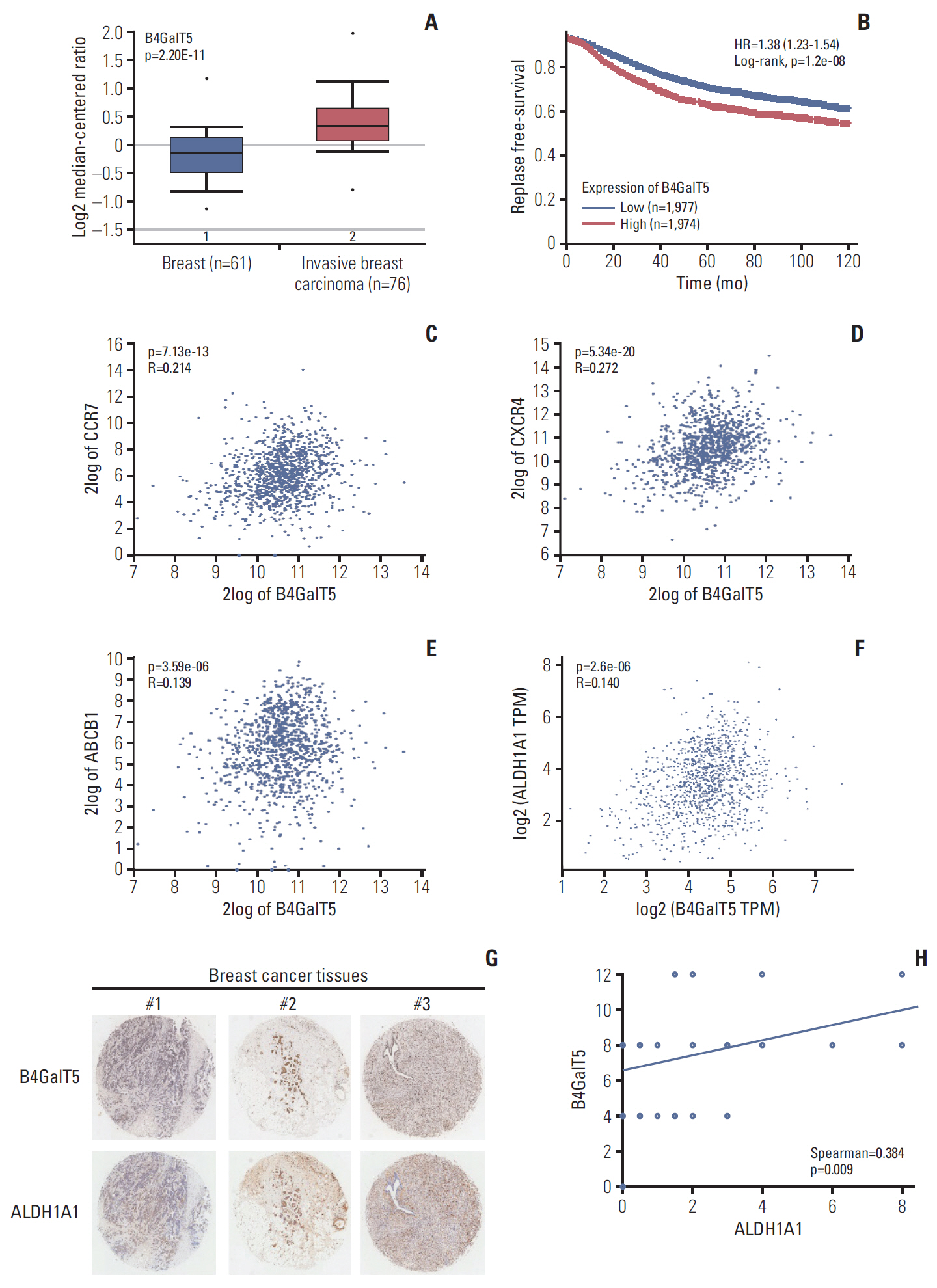
Fig. 1. β1,4-Galactosyltransferase V (B4GalT5) is associated with the stemness of breast cancer. (A) The Cancer Genome Atlas (TCGA) breast dataset analysis of B4GalT5 mRNA levels in invasive breast carcinomas (n=76) and normal breast tissues (n=61) using the Oncomine browser. (B) Kaplan-Meier survival curves for patients with breast cancer (n=3,955) in 120 months. The correlation between B4GalT5 expression and relapse-free survival of patients with breast cancer were analyzed using Kaplan-Meier plotter. (C-E) Correlation between B4GalT5 and CC-chemokine receptor 7 (CCR7) (C), C-X-C chemokine receptor 4 (CXCR4) (D), or ATP binding cassette subfamily B member 1 (ABCB1) (E) mRNA levels in invasive breast carcinoma samples from TCGA dataset (n=1,097) using ‘R2: Genomics Analysis and Visualization Platform.’ (F) The correlation between B4GalT5 and aldehyde dehydrogenase 1A1 (ALDH1A1) mRNA levels in breast carcinoma patients from TCGA dataset using GEPIA 2. (G) Representative images of B4GalT5 and ALDH1A1 expression in the breast tumor microarray by immunohistochemistry (IHC) analysis. (H) The correlation between B4GalT5 and ALDH1A1 IHC scores of 45 cores of the breast tumor microarray.
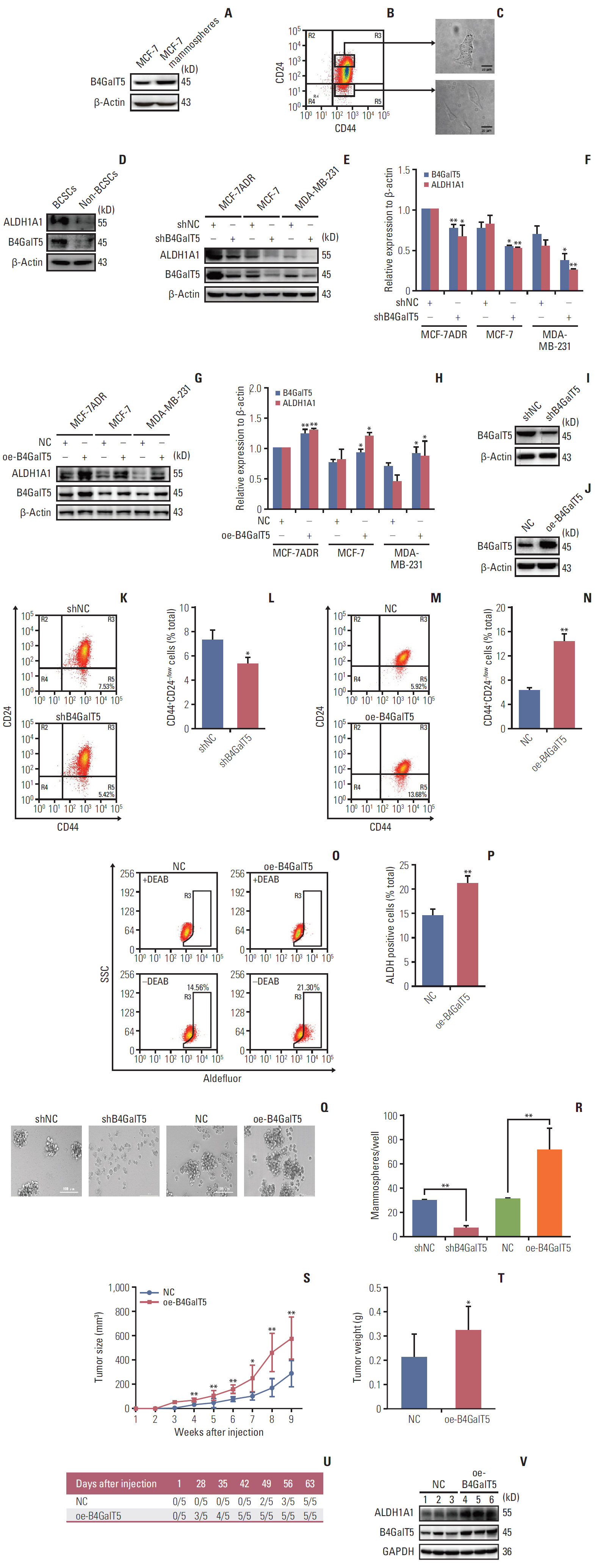
Fig. 2. β1,4-Galactosyltransferase V (B4GalT5) is required for maintaining the stemness of breast cancer. (A) Western blotting analysis of B4GalT5 expression levels in intact MCF-7 cells and MCF-7 mammospheres. (B) Flow cytometry (FCM) analysis and sorting of CD44+CD24–/low cells (breast cancer stem cells [BCSCs]) and CD44+CD24+ cells (non-BCSCs) in MCF-7ADR cells. (C) Patterns of BCSCs and non-BCSCs sorted from MCF-7ADR cells. Scale bars=20 μm. (D) Western blotting analysis of B4GalT5 and aldehyde dehydrogenase 1A1 (ALDH1A1) expression levels in BCSCs and non-BCSCs sorted from MCF-7ADR cells. (E-H) Effects of B4GalT5 on ALDH1A1 expression in breast cancer cells by western blotting. MCF-7, MCF-7ADR, and MDA-MB-231 cells were transfected using shB4GalT5 (E, F) or B4GalT5 (G, H) plasmids for 48 hours and subjected to western blotting compared to the negative control. Protein band densities were quantified by normalizing to β-actin. (I, J) Expression levels of B4GalT5 in MCF-7ADR/shB4GalT5 (I) and MCF-7ADR/oe-B4GalT5 (J) compared to the negative control. (K-N) Influences of B4GalT5 on the proportion of CD44+CD24–/low cells in MCF-7ADR/shB4GalT5 (K, L) or MCF-7ADR/oe-B4GalT5 (M, N) cells compared to the negative control. Cells were stained with anti-CD44 and anti-CD24 antibodies and analyzed by FCM. (O, P) FCM analysis of ALDH+ cells in MCF-7ADR/oe-B4GalT5 cells compared to MCF-7ADR/NC cells. The cells were stained and analyzed as described. (Q, R) The sizes and numbers of mammospheres formed by MCF-7ADR/shB4GalT5 and MCF-7ADR/oe-B4GalT5 compared to the negative control groups. Two thousand cells per well were cultured in serum-free medium for seven days. Formed mammospheres were quantified, and images of formed spheres with a diameter of > 50 μm were taken with a light microscope. Scale bars=100 μm. (S, T) Volumes (S) and weights (T) of tumor formed by MCF-7ADR/NC and MCF-7ADR/oe-B4GalT5 cells in BALB/c nude mice. MCF-7ADR/NC and MCF-7ADR/oe-B4GalT5 cells were subcutaneously transplanted into female BALB/c nude mice at the concentration of 2×106 cells per site. After 9 weeks, the mice were sacrificed for measurement of tumor weights (n=5, per group). (U) Statistics of tumors with a volume of 100 mm3 after subcutaneous injection of MCF-7ADR cells. (V) Western blotting analysis of B4GalT5 and ALDH1A1 expression in tumor tissues. All experiments were performed in three replicates. All data are representative and shown as means±standard error of the mean. *p < 0.05, **p < 0.01, vs. negative control (NC).
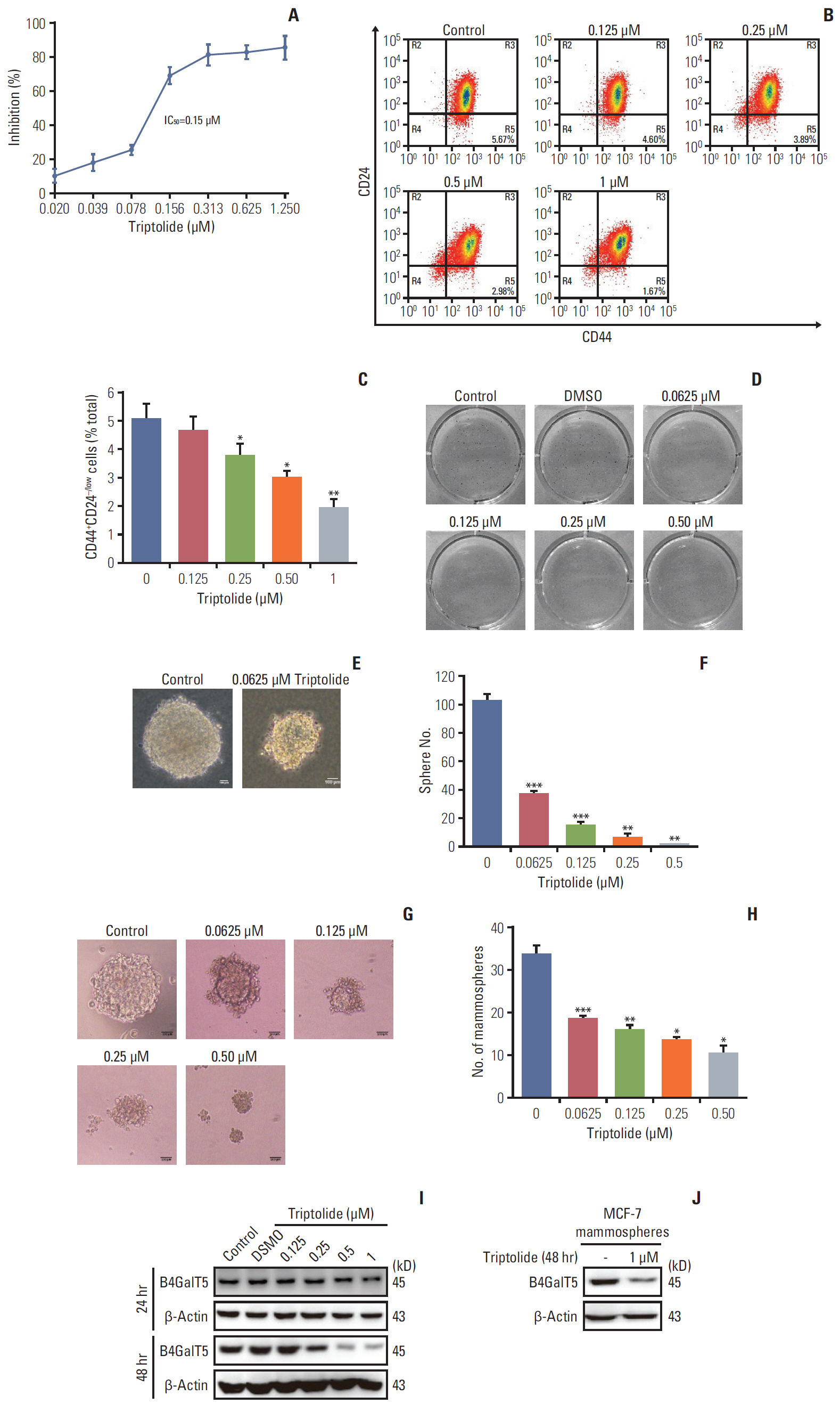
Fig. 3. β1,4-Galactosyltransferase V (B4GalT5) is involved in the inhibition of breast cancer stem cells by triptolide. (A) IC50 of triptolide against MCF-7ADR cells using MTT assay. Cells were treated with increasing concentrations of triptolide for 72 hours and MTT assay was performed. NC, normal control. (B, C) Inhibitory effect of triptolide on the proportion of CD44+CD24–/low cells in MCF-7ADR cells. Cells were treated with increasing concentrations of triptolide for 48 hours, then stained with anti-CD44 and anti-CD24 antibodies and analyzed by flow cytometry. (D) Images of colonies in the soft agar. Five hundred MCF-7ADR cells per well were plated in the upper agar for 30 days with the treatment with triptolide before MTT dye was added. Images of formed colonies containing up to 50 cells with a light microscope are shown. (E) Images of colonies magnified treated with dimethyl sulfoxide (DMSO) or 0.0625 μM triptolide using a light microscope. Scale bars=100 μm. (F) Quantification of the number of colonies. (G) Images of mammospheres in serum-free medium. 2000 MCF-7 cells per well were cultured in serum-free medium with treatment with phosphate buffered saline (PBS) or different concentrations of triptolide. After 7 days, pictures were taken using light microscopy with a camera. Scale bars=200 μm. (H) Quantification of the number of colonies. (I) Inhibitory effect of triptolide on B4GalT5 expression in MCF-7ADR cells in dose- and time-dependent manners. MCF-7ADR cells were treated with different concentrations of triptolide for 24 hours and 48 hours. Then cells were subjected to western blotting. (J) Inhibitory effect of triptolide on B4GalT5 expression in MCF-7 mammospheres by western blotting. All experiments were performed in three replicates. All data are representative and shown as means±standard error of the mean. *p < 0.05, **p < 0.01, ***p < 0.001, vs. the former group.
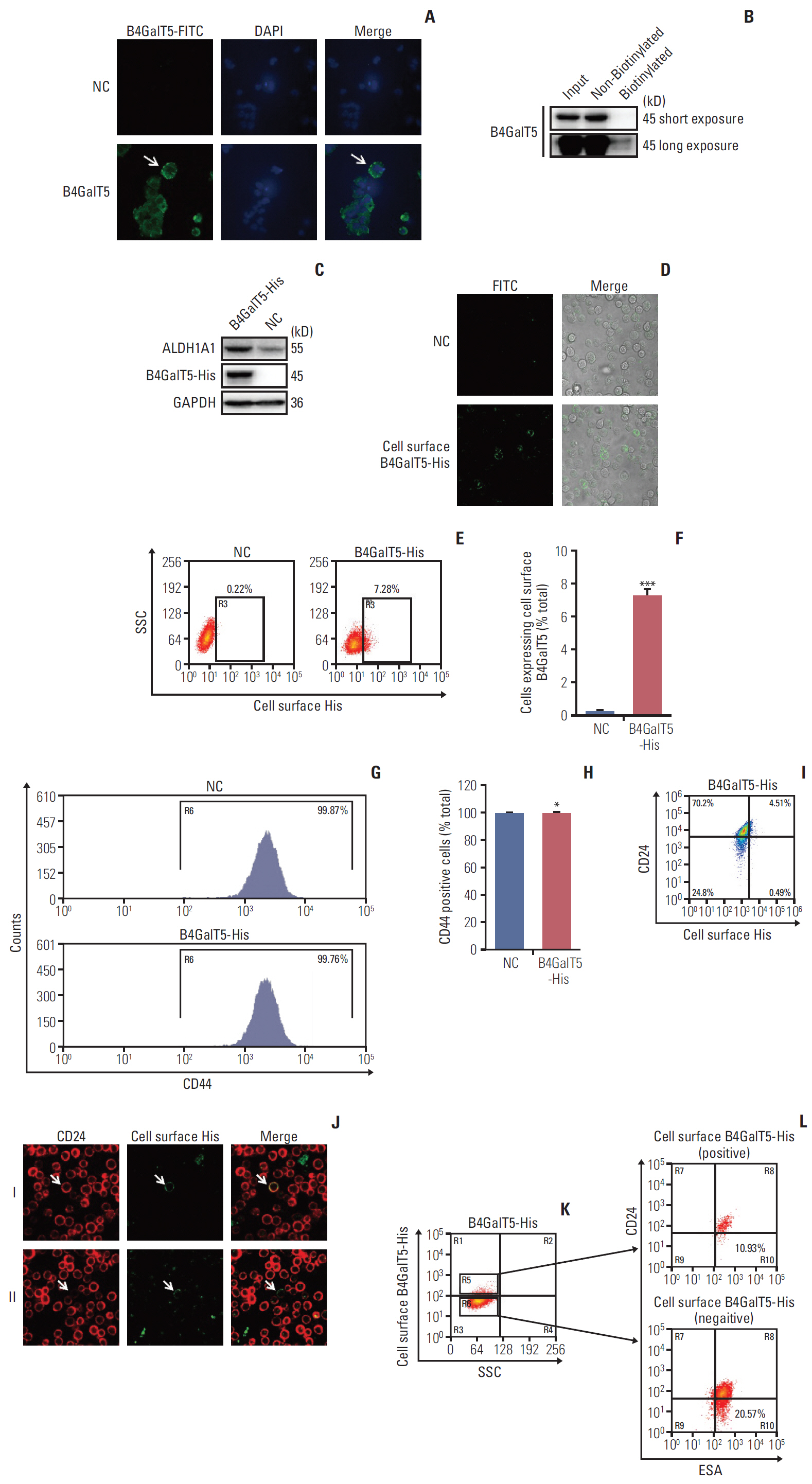
Fig. 4. Cell surface β1,4-galactosyltransferase V (B4GalT5) is not responsible for the stemness of breast cancer. (A) Immunofluorescence analysis of B4GalT5 expression in MCF-7ADR cells. Cells were stained with IgG or anti-B4GalT5 antibody followed by fluorescein isothiocyanate (FITC)–conjugated anti-Rabbit antibodies as described and analyzed by a laser scanning confocal microscope. (B) Cell surface biotinylation assay to compare B4GalT5 localization in plasma membrane and cytoplasmic fractions of MCF-7ADR cells. (C) Construction of MCF-7ADR cells that stably expresses B4GalT5 with a C-terminal 6×His tag. MCF-7ADR cells were transfected with pBABE-B4GalT5-His-IRES-puro plasmid for 48 hours and then selected with 150 μg/mL puromycin. Expression of corresponding proteins was examined by western blotting. (D) Representative images of B4GalT5-His expression on the cell surface of MCF-7ADR/B4GalT5-His and MCF-7ADR/NC cells. After digested and resuspended, the cells were stained with anti-His antibody at room temperature for 1 hour followed by FITC-conjugated anti-rabbit antibody at room temperature for 1 hour. Pictures were taken by a laser scanning confocal microscope. (E, F) Flow cytometry (FCM) analysis of B4GalT5-His expression on the cell surface of MCF-7ADR/B4GalT5-His and MCF-7ADR/NC cells. Cells were stained with phycoerythrin (PE)-conjugated anti-His antibody at room temperature for 1 hour and analyzed by Moflo XDP. Error bars represent standard error of the mean (n=3, ***p < 0.001, t test). (G, H) FCM analysis of CD44 expression in MCF-7ADR/B4GalT5-His and MCF-7ADR/NC cells. The cells were stained with FITC-conjugated CD44 antibody at 4℃ for 30 minutes and analyzed using Moflo XDP. Error bars represent SEM (n=3; *p > 0.05, t test). (I) FCM analysis of CD24–/low cells in B4GalT5-His positive on the cell surface of MCF-7ADR/B4GalT5-His cells. MCF-7ADR/B4GalT5-His and MCF-7ADR/NC cells were stained with APC-conjugated anti-CD24 and PE-conjugated anti-His antibodies and analyzed by Moflo XDP. (J) Representative images of CD24 and B4GalT5-His expression on the cell surface of MCF-7ADR/B4GalT5-His and MCF-7ADR/NC cells. After digested and resuspended, the cells were stained with anti-CD24 and anti-His antibodies. Pictures were taken by a laser scanning confocal microscope. (K, L) FCM analysis of ESA+CD24–/low cells in B4GalT5-His positive (K, R5) and negative (K, R6) on the cell surface of MCF-7ADR/B4GalT5-His cells; NC, negative control; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; SSC, Side Scatter; ESA, epithelial specific antigen. All experiments were performed in three replicates.
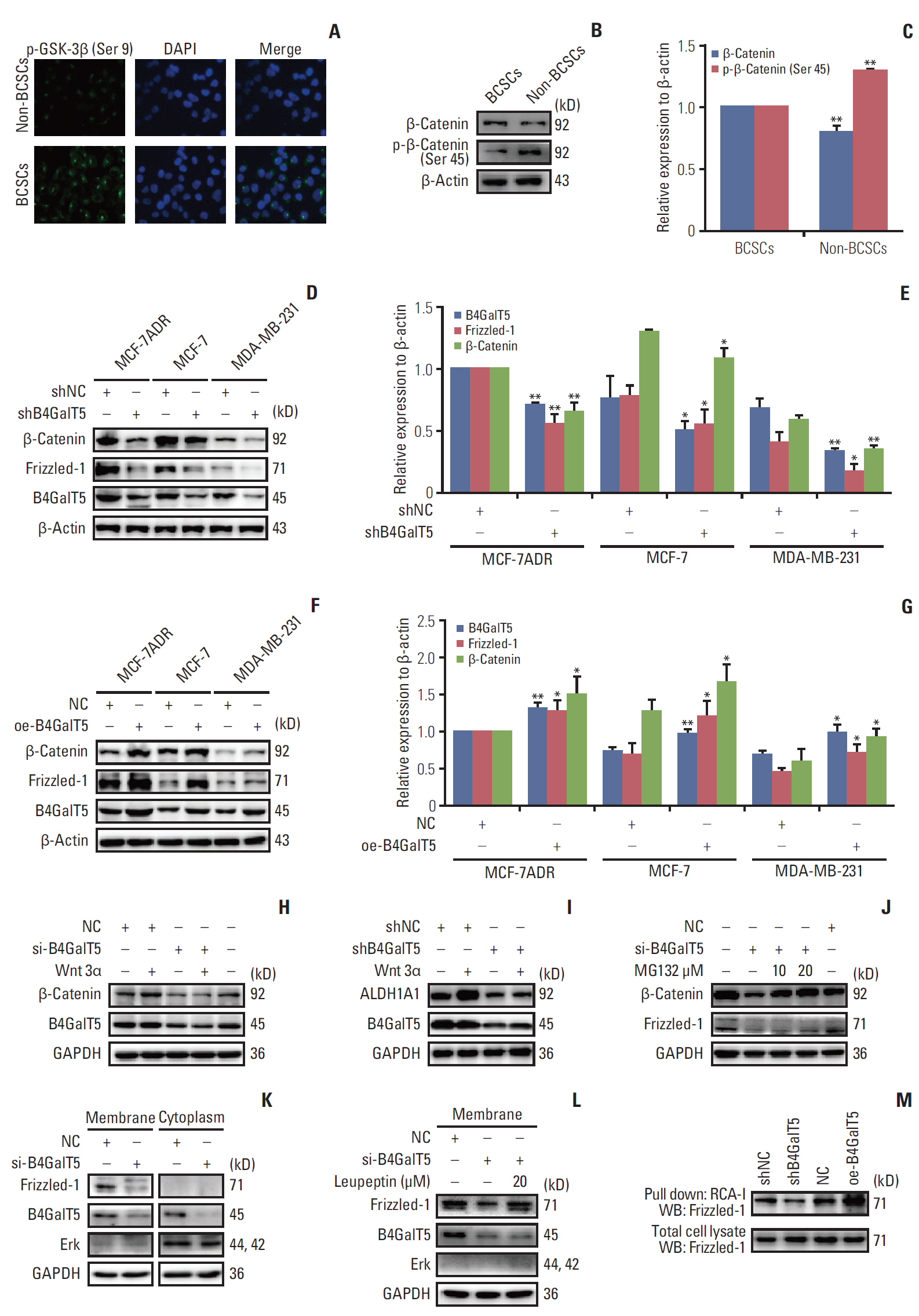
Fig. 5. β1,4-Galactosyltransferase V (B4GalT5) promotes Wnt/β-catenin signaling that is hyperactivated in breast cancer stem cells (BCSCs). (A) Immunofluorescence analysis of phosphorylated-GSK3β (Ser 9) expression in BCSCs and non-BCSCs. MCF-7ADR cells were sorted into BCSCs and non-BCSCs, then stained with antibodies and DAPI as described and analyzed by a laser scanning confocal microscope. (B, C) Western blotting analysis of β-catenin and phosphorylated-β-catenin (Ser 45) expression levels in BCSCs and non-BCSCs. Protein band densities were quantified by normalizing to β-actin. (D, E) The effect of B4GalT5 knockdown on Frizzled-1, β-catenin, and B4GalT5 expression levels in MCF-7ADR, MCF-7, and MDA-MB-231 cell lines by western blotting analysis. Protein band densities were quantified by normalizing to β-actin. (F, G) The effect of B4GalT5 overexpression on Frizzled-1, β-catenin, and B4GalT5 expression levels in MCF-7ADR, MCF-7, and MDA-MB-231 cell lines by western blotting analysis. Protein band densities were quantified by normalizing to β-Actin. (H) The effect of B4GalT5 on wnt 3α induced Wnt/β-catenin signaling by western blotting (WB). After transfected using B4GalT5 siRNA for 12 hours, MCF-7ADR cells were starved for 36 hours followed by adding wnt 3α, and subjected to western blotting. (I) The effect of B4GalT5 on Wnt 3α induced ALDH1A1 expression by western blotting. MCF-7ADR/shNC and MCF-7ADR/shB4GalT5 cells were starved for 36 hours followed by adding Wnt 3α, and subjected to western blotting. (J) The degradation pathway of β-catenin due to B4GalT5 knockdown. After transfected using B4GalT5 siRNA for 41 hours, MCF-7ADR cells were treated with MG132 (protease inhibitor) for 7 hours. Protein levels were examined by western blotting. (K) Western blotting analysis of membrane and cytosol proteins in MCF-7ADR cells. MCF-7ADR cells were transfected using B4GalT5 siRNA for 48 hours, extracted into membrane and cytosol proteins and then subjected to western blotting. Erk was used for examining purity of membrane proteins. (L) The degradation pathway of Frizzled-1 on the cell surface of MCF-7ADR cells due to B4GalT5 knockdown. After transfected using B4GalT5 siRNA for 40 hours, MCF-7ADR cells were treated with 20 μM leupeptin (lysosome inhibitor) for 8 hours. The membrane proteins were extracted as described and then subjected to western blotting. Erk was used for examining purity of membrane proteins. (M) The effect of B4GalT5 on biosynthesis of galactosyl-oligosaccharides in glycans of Frizzled-1 by RCA-I lectin pull-down assay. MCF-7ADR/shNC, MCF-7ADR/shB4GalT5, MCF-7ADR/NC, and MCF-7ADR/oe-B4GalT5 cells were lysed and incubated with agarose-bound RCA-I overnight. Total Frizzled-1 was used as loading control. All experiments were performed in three replicates. Data are presented as means±standard error of the mean. *p < 0.05, **p < 0.01 versus the control group.
Reference
-
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018; 68:394–424.
Article2. Visvader JE, Lindeman GJ. Cancer stem cells: current status and evolving complexities. Cell Stem Cell. 2012; 10:717–28.
Article3. Ghasemi F, Sarabi PZ, Athari SS, Esmaeilzadeh A. Therapeutics strategies against cancer stem cell in breast cancer. Int J Biochem Cell Biol. 2019; 109:76–81.
Article4. Fuster MM, Esko JD. The sweet and sour of cancer: glycans as novel therapeutic targets. Nat Rev Cancer. 2005; 5:526–42.
Article5. Pinho SS, Reis CA. Glycosylation in cancer: mechanisms and clinical implications. Nat Rev Cancer. 2015; 15:540–55.
Article6. Wassler M. β1,4-galactosyltransferases, potential modifiers of stem cell pluripotency and differentiation. In : Bhartiya D, Lenka N, editors. Pluripotent stem cells. Rijeka: InTech;2013. p. 345–72.7. Ma Y, Zhang P, Wang F, Yang J, Yang Z, Qin H. The relationship between early embryo development and tumourigenesis. J Cell Mol Med. 2010; 14:2697–701.
Article8. Gyorffy B, Lanczky A, Eklund AC, Denkert C, Budczies J, Li Q, et al. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast Cancer Res Treat. 2010; 123:725–31.
Article9. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017; 45:W98–W102.
Article10. Muller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, et al. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001; 410:50–6.
Article11. Robey RW, Pluchino KM, Hall MD, Fojo AT, Bates SE, Gottesman MM. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat Rev Cancer. 2018; 18:452–64.
Article12. Ciccone V, Terzuoli E, Donnini S, Giachetti A, Morbidelli L, Ziche M. Stemness marker ALDH1A1 promotes tumor angiogenesis via retinoic acid/HIF-1alpha/VEGF signalling in MCF-7 breast cancer cells. J Exp Clin Cancer Res. 2018; 37:311.
Article13. Li Y, Xian M, Yang B, Ying M, He Q. Inhibition of KLF4 by statins reverses adriamycin-induced metastasis and cancer stemness in osteosarcoma cells. Stem Cell Reports. 2017; 8:1617–29.14. Yang A, Qin S, Schulte BA, Ethier SP, Tew KD, Wang GY. MYC inhibition depletes cancer stem-like cells in triple-negative breast cancer. Cancer Res. 2017; 77:6641–50.
Article15. Youakim A, Dubois DH, Shur BD. Localization of the long form of beta-1,4-galactosyltransferase to the plasma membrane and Golgi complex of 3T3 and F9 cells by immunofluorescence confocal microscopy. Proc Natl Acad Sci U S A. 1994; 91:10913–7.
Article16. Lim SK, Lu SY, Kang SA, Tan HJ, Li Z, Adrian Wee ZN, et al. Wnt signaling promotes breast cancer by blocking ITCHmediated degradation of YAP/TAZ transcriptional coactivator WBP2. Cancer Res. 2016; 76:6278–89.
Article17. Chung J, Karkhanis V, Baiocchi RA, Sif S. Protein arginine methyltransferase 5 (PRMT5) promotes survival of lymphoma cells via activation of WNT/beta-catenin and AKT/GSK3beta proliferative signaling. J Biol Chem. 2019; 294:7692–710.18. Rada P, Rojo AI, Offergeld A, Feng GJ, Velasco-Martin JP, Gonzalez-Sancho JM, et al. WNT-3A regulates an Axin1/NRF2 complex that regulates antioxidant metabolism in hepatocytes. Antioxid Redox Signal. 2015; 22:555–71.
Article19. Cooper GM. The cell: a molecular approach. 2nd ed. Sunderland, MA: Sinauer Associates Inc.;2000.20. Sola RJ, Griebenow K. Effects of glycosylation on the stability of protein pharmaceuticals. J Pharm Sci. 2009; 98:1223–45.21. Sasaki N, Manya H, Okubo R, Kobayashi K, Ishida H, Toda T, et al. beta4GalT-II is a key regulator of glycosylation of the proteins involved in neuronal development. Biochem Biophys Res Commun. 2005; 333:131–7.22. Chen WS, Chang HY, Li CP, Liu JM, Huang TS. Tumor beta-1,4-galactosyltransferase IV overexpression is closely associated with colorectal cancer metastasis and poor prognosis. Clin Cancer Res. 2005; 11(24 Pt 1):8615–22.23. Yoshihara T, Satake H, Nishie T, Okino N, Hatta T, Otani H, et al. Lactosylceramide synthases encoded by B4galt5 and 6 genes are pivotal for neuronal generation and myelin formation in mice. PLoS Genet. 2018; 14:e1007545.
Article24. Talhaoui I, Bui C, Oriol R, Mulliert G, Gulberti S, Netter P, et al. Identification of key functional residues in the active site of human {beta}1,4-galactosyltransferase 7: a major enzyme in the glycosaminoglycan synthesis pathway. J Biol Chem. 2010; 285:37342–58.25. Katoh M. Canonical and non-canonical WNT signaling in cancer stem cells and their niches: cellular heterogeneity, omics reprogramming, targeted therapy and tumor plasticity (Review). Int J Oncol. 2017; 51:1357–69.
Article26. Wang H, Zhou T, Peng J, Xu P, Dong N, Chen S, et al. Distinct roles of N-glycosylation at different sites of corin in cell membrane targeting and ectodomain shedding. J Biol Chem. 2015; 290:1654–63.
Article27. Li CW, Lim SO, Xia W, Lee HH, Chan LC, Kuo CW, et al. Glycosylation and stabilization of programmed death ligand-1 suppresses T-cell activity. Nat Commun. 2016; 7:12632.
Article28. Xu Y, Chang R, Xu F, Gao Y, Yang F, Wang C, et al. N-glycosylation at Asn 402 stabilizes N-cadherin and promotes cellcell adhesion of glioma cells. J Cell Biochem. 2017; 118:1423–31.
Article29. Yonamine I, Bamba T, Nirala NK, Jesmin N, KosakowskaCholody T, Nagashima K, et al. Sphingosine kinases and their metabolites modulate endolysosomal trafficking in photoreceptors. J Cell Biol. 2011; 192:557–67.
Article30. Rodeheffer C, Shur BD. Targeted mutations in beta1,4-galactosyltransferase I reveal its multiple cellular functions. Biochim Biophys Acta. 2002; 1573:258–70.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Transforming Growth Factor-β Signaling Inhibits the Osteogenic Differentiation of Mesenchymal Stem Cells via Activation of Wnt/β-Catenin Pathway
- Natural Products Targeting Wnt/β-catenin Signaling Pathway
- RNF43 and ZNRF3 in Wnt Signaling - A Master Regulator at the Membrane
- Kinesin Family Member 11 Enhances the Self-Renewal Ability of Breast Cancer Cells by Participating in the Wnt/β-Catenin Pathway
- The crosstalk of Wnt/β-catenin signaling and p53 in acute kidney injury and chronic kidney disease
