G Protein–Coupled Receptor 30 Mediates the Anticancer Effects Induced by Eicosapentaenoic Acid in Ovarian Cancer Cells
- Affiliations
-
- 1State Key Laboratory of Pharmaceutical Biotechnology and Jiangsu Key Laboratory of Molecular Medicine, Medical School of Nanjing University, Nanjing, China
- 2Department of Obstetrics and Gynecology, Xiamen Chang Gung Hospital, Xiamen, China
- 3Obstetrics and Gynecology Hospital, Fudan University, Shanghai, China
- 4Department of Pathology, Nanjing Drum Tower Hospital, The Affiliated Hospital of Nanjing University Medical School, Nanjing, China
- 5Biology Science Institutes, Chongqing Medical University, Yuzhong, China
- KMID: 2504462
- DOI: http://doi.org/10.4143/crt.2019.380
Abstract
- Purpose
While numerous epidemiological studies have indicated that omega-3 polyunsaturated fatty acids have anticancer properties in various cancers, the effects and mechanisms of eicosapentaenoic acid (EPA) in ovarian cancer cell growth are poorly understood.
Materials and Methods
ES2 ovarian clear cell carcinoma cells and SKOV3 adenocarcinoma cells were treated with palmitic acid or EPA, followed by flow cytometry and cell counting to measure apoptosis and proliferation, respectively. A modified protein lipid overlay assay was used to further verify whether EPA was a ligand of G protein–coupled receptor 30 (GPR30) in ES2 cells. The levels of apoptosis-related genes, phosphorylated AKT, and phosphorylated ERK1/2 were detected to explore the underlying mechanism. Finally, inhibitory effect of EPA on tumor growth via GPR30 was determined in vitro and in vivo.
Results
EPA suppressed ES2 ovarian clear cell carcinoma cells growth via GPR30, a novel EPA receptor, by inducing apoptosis. As a ligand of GPR30, EPA activated the GPR30-cAMP– protein kinase A signaling pathway. When GPR30 was suppressed by siRNA or its inhibitor G15, the antiproliferative action of EPA was impaired. Furthermore, EPA inhibited tumor growth by blocking the activation of AKT and ERK. In the mouse xenograft model, EPA decreased tumor volume and weight through GPR30 by blocking tumor cell proliferation.
Conclusion
These results confirm that EPA is a tumor suppressor in human ovarian clear cell carcinoma cells and functions through a novel fatty acid receptor, GPR30, indicating a mechanistic linkage between omega-3 fatty acids and cancers.
Figure
-

Fig. 1. Eicosapentaenoic acid (EPA) inhibits cell proliferation in ovarian cancer cells. (A) Dose-response curves of palmitic acid, palmitoleic acid, oleic acid, linoleic acid, arachidonic acid, linolenic acid, and EPA in ES2 cells and SKOV3 cells. (B) The toxicity of palmitic acid and EPA at different concentrations on ES2 and SKOV3 cells. (C) Cell proliferation of ES2 and SKOV3 cells in time course for various fatty acids in ES2 cells and SKOV3 cells were treated with 300 μM fatty acids. (D) Apoptosis by EPA in ES2 cells. The cells were treated with or without 300 μM EPA for 48 hours. Apoptosis was detected by cleaved caspase-3. PI, propidium iodide. (E) The expression of proapoptotic genes and antiapoptotic genes in ES2 cells treated with or without 300 μM EPA for 48 hours was analyzed by quantitative reverse transcription polymerase chain reaction. (F) The toxicity of EPA on normal ovarian epithelial cells (OEC), ES2, and SKOV3 cells. (G) The apoptosis rate of normal OEC, ES2, and SKOV3 cells treated with EPA. Values are presented as mean±standard deviation from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001.
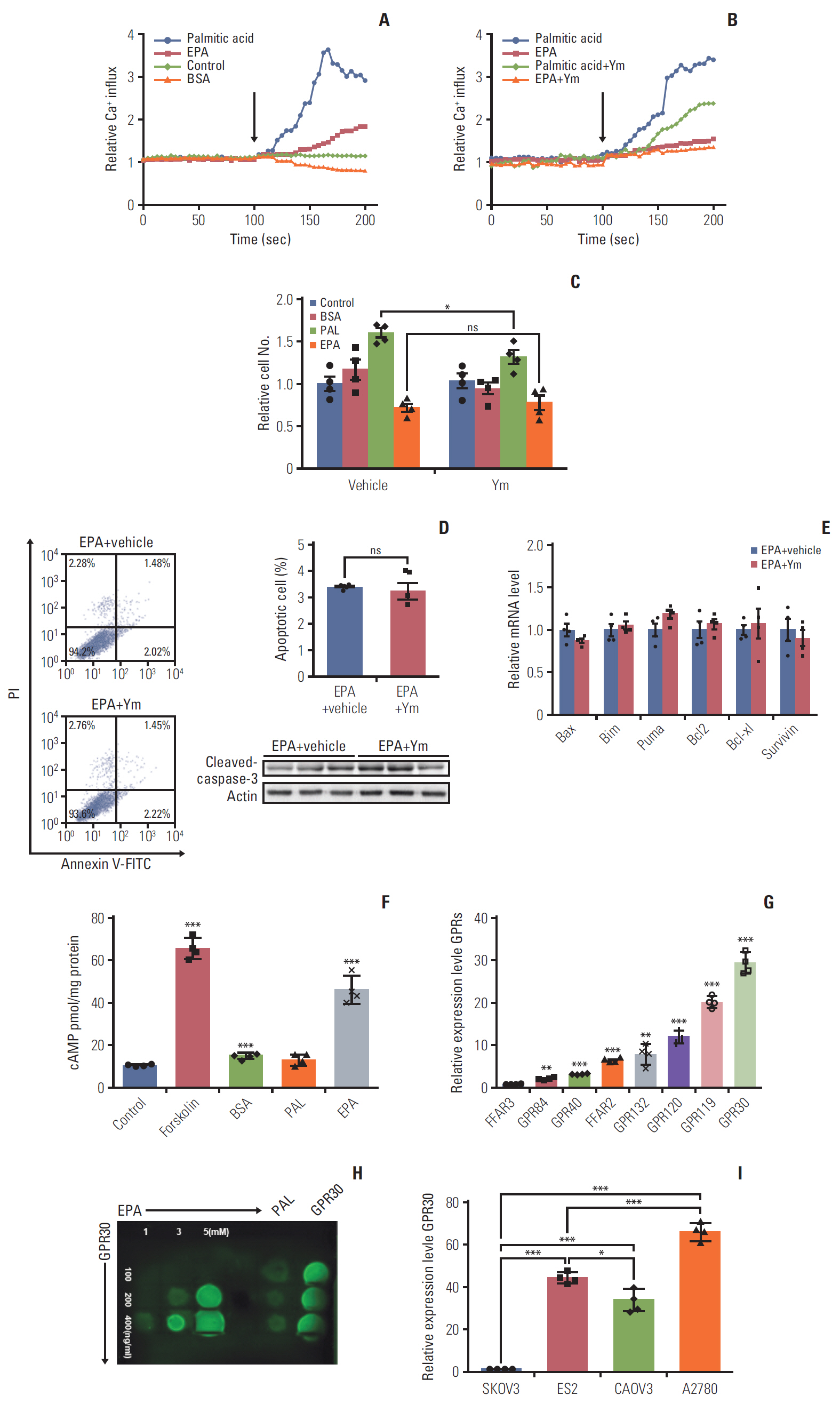
Fig. 2. G protein–coupled receptor 30 (GPR30) acts as a novel eicosapentaenoic acid (EPA) receptor in ES2 cells. (A, B) ES2 cells were loaded with 3 μM of the fluorescent calcium probe fura-2A/M. (A) Cells were stimulated in phosphate buffered saline alone (control), 300 μM palmitic acid bound to bovine serum albumin (BSA) (0.1%), 300 μM EPA bound to BSA (0.1%), or BSA alone. (B) ES2 cells were incubated with YM254890 (Gq protein inhibitor) at 37°C for 5 minutes before the addition of fatty acids. The arrows indicate the onset of stimulation. (C) Cell proliferation of ES2 cells treated with 0.1% BSA, 300 μM palmitic acid (PAL), or 300 μM EPA in the absence or presence of YM254890 for 24 hours. (D) Apoptosis of ES2 cells treated with 300 μM EPA in the absence or presence of YM254890. Apoptosis was detected by cleaved caspase-3. PI, propidium iodide. (E) The expression of proapoptotic genes and antiapoptotic genes in ES2 cells treated with 300 μM EPA in the absence or presence of YM254890 for 48 hours was analyzed by quantitative reverse transcription polymerase chain reaction (RT-PCR). (F) Intracellular cAMP by EPA in ES2 cells. Cells were treated with 0.1% BSA, 300 μM PAL, or 300 μM EPA for 24 hours for cAMP measurement. (G) The relative expression of various GPRs in ES2 cells by RT-PCR. Data were normalized to the expression level of free fatty acid receptor 3 (FFAR3). (H) Interaction of GPR30 and EPA was detected by a protein lipid overlay assay. Increasing amounts of EPA were incubated with increasing concentrations of purified GFP-GPR30 harvested from ES2 cell lysates. (I) Relative expression of GPR30 was measured by quantitative RT-PCR in different ovarian cancer cell lines. Data were normalized to the expression level in SKOV3. Values are presented as mean±standard deviation from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001; ns, not significant.

Fig. 3. G protein–coupled receptor 30 (GPR30) is expressed in patients with various ovarian cancers. The expression level of GPR30 in patients with various ovarian cancers according to a tissue microarray and immunohistochemistry. (A) Immunohistochemical staining of GPR30 in ovarian tissues with various ovarian cancers. (B) The expression grade of GPR30 in patients with various ovarian cancers (n=173). HGSC, high-grade serous carcinoma; LGSC, low-grade serous carcinoma; MC, mucinous carcinoma, CCC, clear cell carcinoma; EC, endometrioid adenocarcinoma. (C) The correlation analysis of GPR30 expression level with overall survival of patients (The grade score 0 to 3 represents the intensity of GPR30 from lowest to highest).
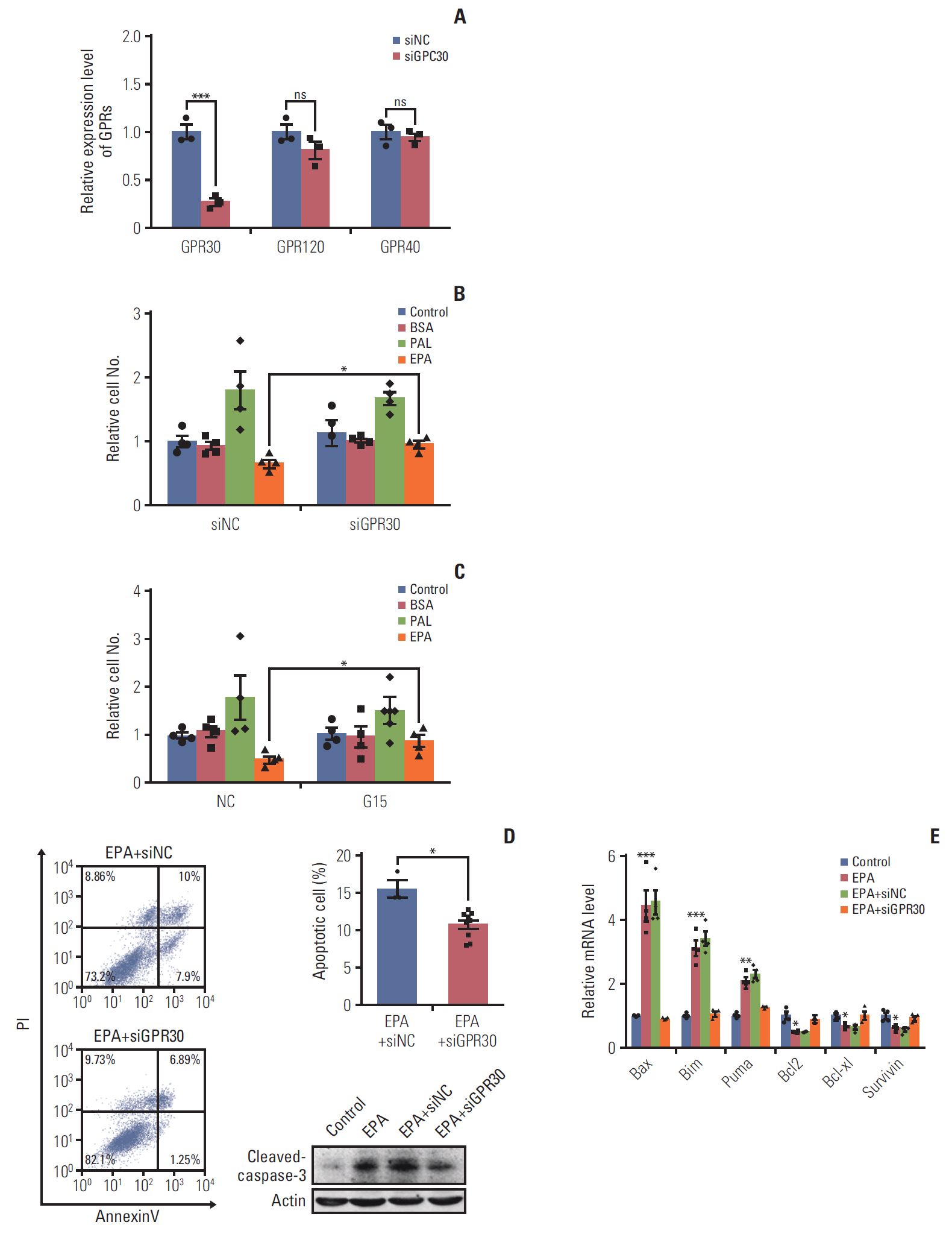
Fig. 4. G protein–coupled receptor 30 (GPR30) is involved in eicosapentaenoic acid (EPA)–induced cell antiproliferation and proapoptosis in ES2 cells. (A) The efficacy of GPR30 small interfering RNA (siRNA). (B) Cell proliferation of ES2 cells treated with 0.1% bovine serum albumin (BSA), 300 μM palmitic acid (PAL), or 300 μM EPA with or without GPR30 siRNA treatment. (C) Cell proliferation of ES2 cells treated with 0.1% BSA, 300 μM PAL, or 300 μM EPA with or without G15. (D) Apoptosis of ES2 cells treated with 300 μM EPA and control RNA interference (RNAi) or 300 μM EPA and GPR30 RNAi for 48 hours. Apoptosis was detected by cleaved caspase-3. PI, propidium iodide. (E) The expression of proapoptotic genes and antiapoptotic genes in ES2 cells treated with 300 μM EPA or 300 μM EPA and GPR30 siRNA for 48 hours was analyzed by quantitative reverse transcription polymerase chain reaction. Values are presented as mean±standard deviation from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001.
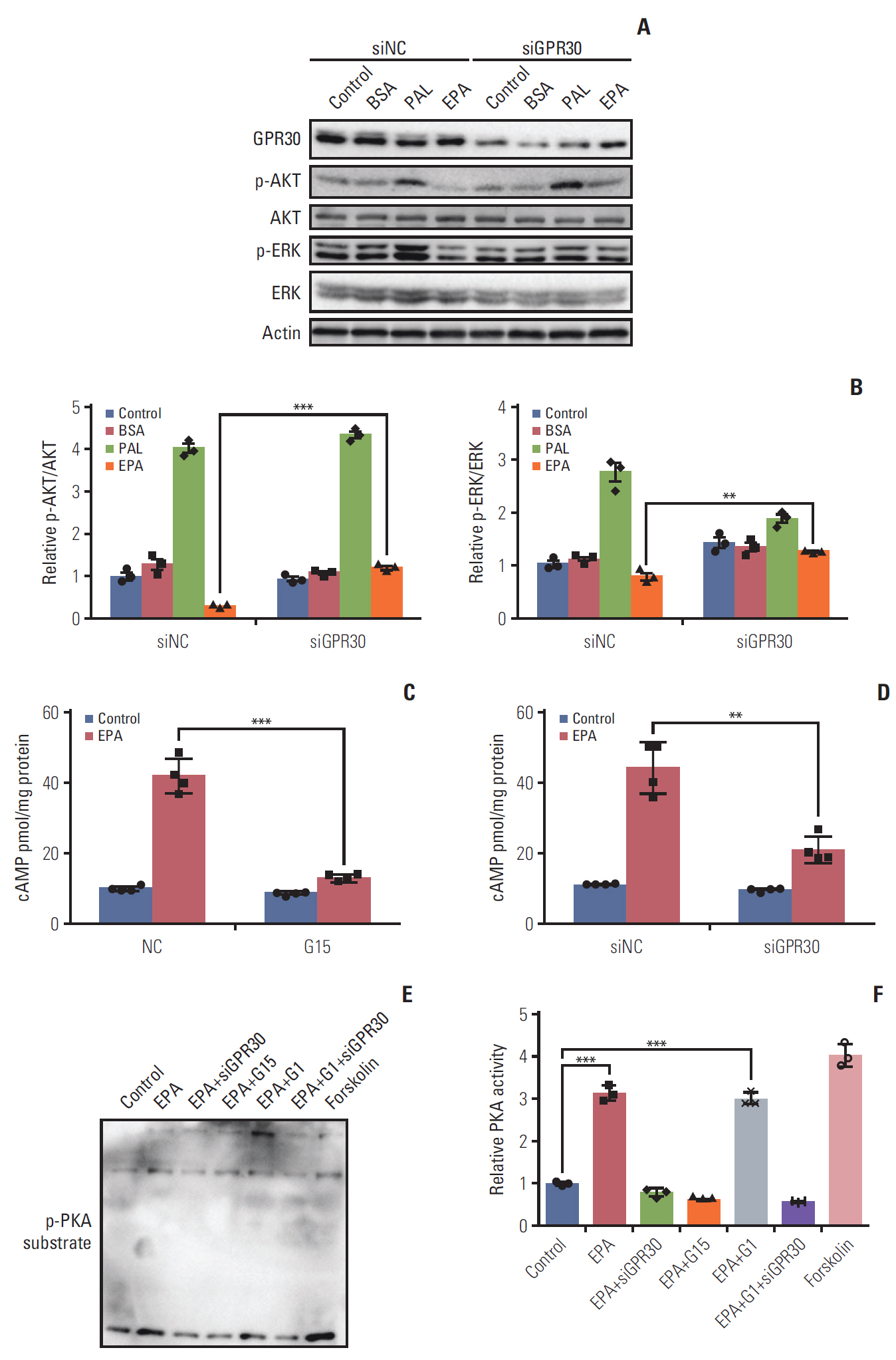
Fig. 5. Eicosapentaenoic acid (EPA) participates in the G protein–coupled receptor 30 (GPR30)–cAMP–protein kinase A signaling pathway in ES2 cells. (A) After 24 hours of serum starvation in Opti-MEM, cells were incubated for 24 hours in OptiMEM and stimulated with phosphate buffered saline alone (control), 0.1% bovine serum albumin (BSA), 300 μM palmitic acid (PAL) bound to BSA (0.1%) or 300 μM EPA bound to BSA (0.1%) with or without GPR30 siRNA treatment. (B) p-AKT/AKT and p-ERK/ERK were analyzed by Western blotting and quantified by ImageJ. (C, D) ES2 cells were treated with or without 300 μM EPA in the absence and presence of G15 (C) or GPR30 siRNA (D) for 24 hours for cAMP measurement. (E) Phospho-(Ser/Thr) substrate was detected by Western blotting in ES2 cells treated with EPA, EPA+GPR30 siRNA, EPA+G15, EPA+G1 (500 nM, the specific agonist of GPR30) or EPA+G1+GPR30 siRNA. (F) PKA activities were quantified by ImageJ, and the data are expressed as PKA activity (n=3). Values are presented as mean±standard deviation from three independent experiments. **p < 0.01, ***p < 0.001.
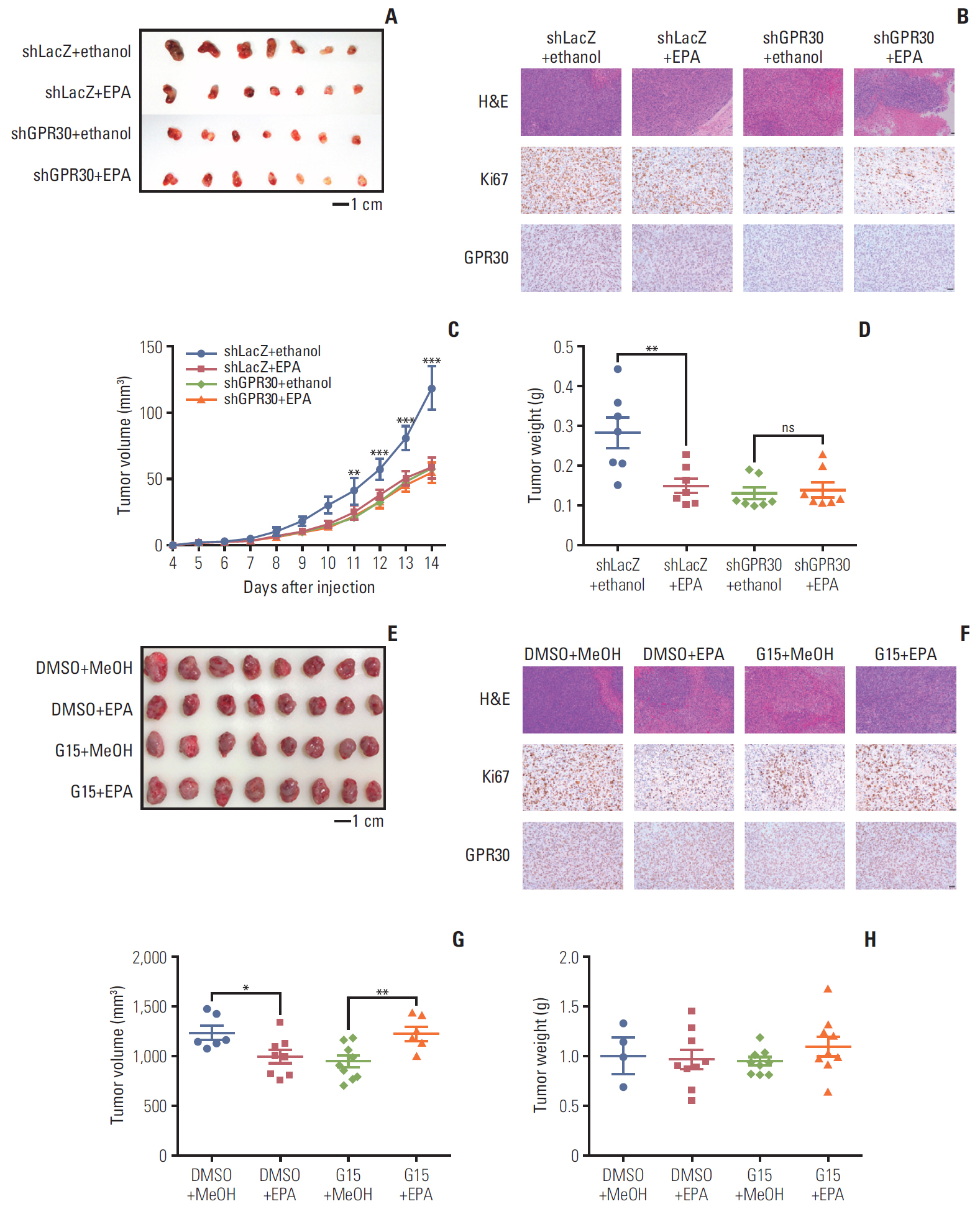
Fig. 6. Eicosapentaenoic acid (EPA) blocks tumor growth via G protein–coupled receptor 30 (GPR30) in mouse xenografts. (A, B) Nude mice bearing ovarian tumors (ES2 cells) were received ethanol in combination with LacZ shRNA as a control, EPA in combination with LacZ shRNA, ethanol in combination with GPR30 shRNA or EPA in combination with GPR30 shRNA. (A) Xenograft tumors (scale bar=1 cm). (B) Ki67 and GPR30 expression (scale bar=50 μm). Tumor volume (C) and tumor weight (D) in (A). (E, F) Nude mice bearing ovarian tumors (ES2 cells) were received dimethyl sulfoxide (DMSO) in combination with MeOH as a control, EPA in combination with DMSO, MeOH in combination with G15 or EPA in combination with G15. (E) Xenograft tumors (scale bar=1 cm). (F) Ki67 and GPR30 expression (scale bar=50 μm). Tumor volume (G) and tumor weight (H) in (E). Values are presented as mean±standard deviation from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001.
Reference
-
References
1. Sapienza C, Issa JP. Diet, nutrition, and cancer epigenetics. Annu Rev Nutr. 2016; 36:665–81.
Article2. Rose DP, Connolly JM. Omega-3 fatty acids as cancer chemopreventive agents. Pharmacol Ther. 1999; 83:217–44.
Article3. Velentzis LS, Keshtgar MR, Woodside JV, Leathem AJ, Titcomb A, Perkins KA, et al. Significant changes in dietary intake and supplement use after breast cancer diagnosis in a UK multicentre study. Breast Cancer Res Treat. 2011; 128:473–82.
Article4. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010; 60:277–300.
Article5. Brinton EA, Mason RP. Prescription omega-3 fatty acid products containing highly purified eicosapentaenoic acid (EPA). Lipids Health Dis. 2017; 16:23.
Article6. Santos CR, Schulze A. Lipid metabolism in cancer. FEBS J. 2012; 279:2610–23.
Article7. Maggiolini M, Picard D. The unfolding stories of GPR30, a new membrane-bound estrogen receptor. J Endocrinol. 2010; 204:105–14.
Article8. Heublein S, Mayr D, Friese K, Jarrin-Franco MC, Lenhard M, Mayerhofer A, et al. The G-protein-coupled estrogen receptor (GPER/GPR30) in ovarian granulosa cell tumors. Int J Mol Sci. 2014; 15:15161–72.
Article9. Mazzuca MQ, Mata KM, Li W, Rangan SS, Khalil RA. Estrogen receptor subtypes mediate distinct microvascular dilation and reduction in [Ca2+]I in mesenteric microvessels of female rat. J Pharmacol Exp Ther. 2015; 352:291–304.
Article10. Jin J, Mao Y, Thomas D, Kim S, Daniel JL, Kunapuli SP. RhoA downstream of G(q) and G(12/13) pathways regulates protease-activated receptor-mediated dense granule release in platelets. Biochem Pharmacol. 2009; 77:835–44.
Article11. Taniguchi M, Nagai K, Arao N, Kawasaki T, Saito T, Moritani Y, et al. YM-254890, a novel platelet aggregation inhibitor produced by Chromobacterium sp. QS3666. J Antibiot (Tokyo). 2003; 56:358–63.
Article12. Wang C, Lv X, Jiang C, Davis JS. The putative G-protein coupled estrogen receptor agonist G-1 suppresses proliferation of ovarian and breast cancer cells in a GPER-independent manner. Am J Transl Res. 2012; 4:390–402.13. Bai LY, Weng JR, Hu JL, Wang D, Sargeant AM, Chiu CF. G15, a GPR30 antagonist, induces apoptosis and autophagy in human oral squamous carcinoma cells. Chem Biol Interact. 2013; 206:375–84.
Article14. Pal D, Dasgupta S, Kundu R, Maitra S, Das G, Mukhopadhyay S, et al. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat Med. 2012; 18:1279–85.
Article15. Hu L, Lau SH, Tzang CH, Wen JM, Wang W, Xie D, et al. Association of Vimentin overexpression and hepatocellular carcinoma metastasis. Oncogene. 2004; 23:298–302.
Article16. Sugiyama T, Kamura T, Kigawa J, Terakawa N, Kikuchi Y, Kita T, et al. Clinical characteristics of clear cell carcinoma of the ovary: a distinct histologic type with poor prognosis and resistance to platinum-based chemotherapy. Cancer. 2000; 88:2584–9.17. Landrum LM, Java J, Mathews CA, Lanneau GS Jr, Copeland LJ, Armstrong DK, et al. Prognostic factors for stage III epithelial ovarian cancer treated with intraperitoneal chemotherapy: a Gynecologic Oncology Group study. Gynecol Oncol. 2013; 130:12–8.
Article18. Kwan HY, Fu X, Liu B, Chao X, Chan CL, Cao H, et al. Subcutaneous adipocytes promote melanoma cell growth by activating the Akt signaling pathway: role of palmitic acid. J Biol Chem. 2014; 289:30525–37.19. Sun H, Hu Y, Gu Z, Owens RT, Chen YQ, Edwards IJ. Omega3 fatty acids induce apoptosis in human breast cancer cells and mouse mammary tissue through syndecan-1 inhibition of the MEK-Erk pathway. Carcinogenesis. 2011; 32:1518–24.
Article20. Kotarsky K, Nilsson NE, Flodgren E, Owman C, Olde B. A human cell surface receptor activated by free fatty acids and thiazolidinedione drugs. Biochem Biophys Res Commun. 2003; 301:406–10.
Article21. Gu Z, Suburu J, Chen H, Chen YQ. Mechanisms of omega-3 polyunsaturated fatty acids in prostate cancer prevention. Biomed Res Int. 2013; 2013:824563.
Article22. Dorsam RT, Gutkind JS. G-protein-coupled receptors and cancer. Nat Rev Cancer. 2007; 7:79–94.
Article23. Prossnitz ER, Oprea TI, Sklar LA, Arterburn JB. The ins and outs of GPR30: a transmembrane estrogen receptor. J Steroid Biochem Mol Biol. 2008; 109:350–3.
Article24. Shu YJ, Weng H, Ye YY, Hu YP, Bao RF, Cao Y, et al. SPOCK1 as a potential cancer prognostic marker promotes the proliferation and metastasis of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol Cancer. 2015; 14:12.
Article25. Brazil DP, Yang ZZ, Hemmings BA. Advances in protein kinase B signalling: AKTion on multiple fronts. Trends Biochem Sci. 2004; 29:233–42.
Article26. Castoria G, Migliaccio A, D'Amato L, Di Stasio R, Ciociola A, Lombardi M, et al. Integrating signals between cAMP and MAPK pathways in breast cancer. Front Biosci. 2008; 13:1318–27.
Article27. Nieman KM, Kenny HA, Penicka CV, Ladanyi A, Buell-Gutbrod R, Zillhardt MR, et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat Med. 2011; 17:1498–503.
Article28. Fukui M, Kang KS, Okada K, Zhu BT. EPA, an omega-3 fatty acid, induces apoptosis in human pancreatic cancer cells: role of ROS accumulation, caspase-8 activation, and autophagy induction. J Cell Biochem. 2013; 114:192–203.
Article29. Sabharwal SS, Schumacker PT. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles' heel? Nat Rev Cancer. 2014; 14:709–21.
Article30. Gajate C, Gonzalez-Camacho F, Mollinedo F. Lipid raft connection between extrinsic and intrinsic apoptotic pathways. Biochem Biophys Res Commun. 2009; 380:780–4.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- 1,3,5-Tricaffeoylquinic Acid from Ipomoea batatas Vines Induced Ovarian Cancer Cell Apoptosis and Inhibited Endothelial Tube Formation
- Stimulatory Anticancer Effect of Resveratrol Mediated by G Protein-Coupled Estrogen Receptor in Colorectal Cancer
- Action and Signaling of Lysophosphatidylethanolamine in MDA-MB-231 Breast Cancer Cells
- Growth Regulation of Ovarian Cancer Cells through the Inactivation of AP-1 by Retinoid Derivatives
- Differential Gene Expression in GPR40-Overexpressing Pancreatic beta-cells Treated with Linoleic Acid
