Bovine adenovirus type 3 virions cannot be rescued in vivo after full-length viral genome transfection in the absence of detectable polypeptide IX
- Affiliations
-
- 1College of Veterinary Medicine, North-west A&F University, Yangling 712100, China. duenqi227@126.com
- 2Chinese Institute of Veterinary Drug Controls, Beijing 100000, China.
- 3VIDO-InteVac, School of Public Health, University of Saskatchewan Saskatoon, Saskatchewan S4S 6A6, Canada. duenqi227@126.com
- 4Vaccinology & Immunotherapeutics Program, School of Public Health, University of Saskatchewan Saskatoon, Saskatchewan S4S 6A6, Canada.
- KMID: 2412575
- DOI: http://doi.org/10.4142/jvs.2017.18.2.217
Abstract
- Bovine adenovirus type 3 (BAdV3) is being used in the development of potential vehicles for gene therapy and vectored vaccine. To that end, a more comprehensive description of BAdV3 biology is essential. In this study, we focused on the role of pIX in BAdV3 virion rescue after full-length BAdV3 genome transfection. Initially, pIX deletion or initiation codon mutation abolished the production of progeny virions, which suggested that pIX was essential for the rescue of BAdV3 containing a full-length genome. Moreover, through transfection of a panel of pIX mutant BAdV3 genomes, we observed that the conserved N-terminus and the putative leucine zipper element (PLZP) were essential for virion rescue, whereas the C-terminus following the coiled-coil domain was non-essential. In addition, swap of the PLZP element and its following region of BAdV3 pIX to corresponding domains of human adenovirus type 5 (HAdV5) did not affect virion production, whereas swap of the entire pIX abolished production of progeny virions. We suggest that failure of the full-length BAdV3 pIX swap might be due to species specificity of its N-terminus region before the PLZP element.
MeSH Terms
Figure
-
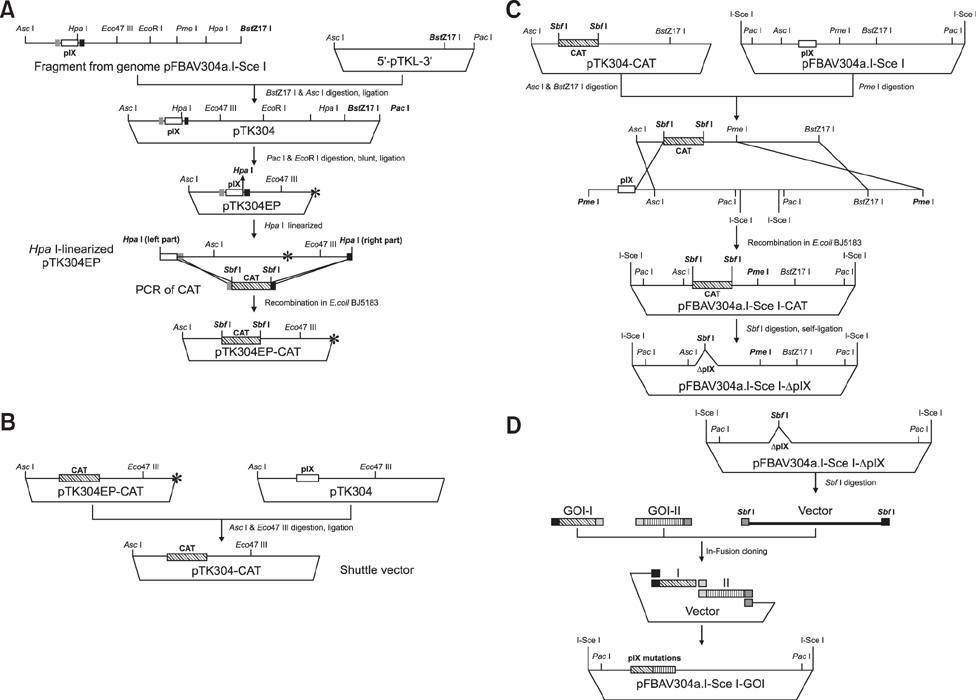
Fig. 1 Strategy used for the generation of recombinant BAdV3 plasmids. (A) The flowchart for construction of plasmid pTK304EP-chloramphenicol acetyltransferase (CAT). Firstly, a 12176 AscI-BstZ17I DNA fragment (nucleotide [nt] 1403 to nt 13578) from pFBAV304a.I-SceI was ligated into AscI-BstZ17I- cleaved 5'- pTKL- 3' to create pTK304. Then, digested plasmid pTK304 with EcoRI and PacI was blunted and self-ligated for eliminating the extra HpaI site to generate pTK304EP. Finally, to construct plasmid pTK304EP-CAT gene, a 1040 bp CAT fragment (containing 50 bp homology arms in 5' and 3' sites and flanked with SbfI enzyme sites) was inserted into HpaI-linearized pTKEP304 in Escherichia coil BJ5185 by homology recombination. (B) The flowchart for construction of shuttle plasmid pTK304-CAT. A 5385 bp fragment isolated from pTK304EP-CAT using AscI and Eco47III was ligated into AscI-Eco47III-cleaved pTK304 to created final shuttle plasmid pTK304-CAT. (C) The flowchart for construction of pIX deleted BAdV3 plasmid pFBAV304a. I-SceI-ΔpIX. AscI-BstZ17I-cleaved pTK304-CAT was cloned into PmeI-linearized pFBAV304a.I-SceI by homology recombination in Escherichia coil BJ5185 to construct pFBAV304a.I-SceI-CAT. Then, the pIX deleted recombination plasmid pFBAV304a.I-SceI-ΔpIX was constructed by digesting the recombinant pFBAV304a.I-SceI-CAT with SbfI and religated to release CAT expression cassette. (D) The flowchart for construction of recombinant BAdV3s expressing genes of variant mutated pIXs. The variant pIX mutations were amplified by polymerase chain reaction using primers and templates (Table 1), then inserted in SbfI site of pFBAV304a.I-SceI-ΔpIX using In-Fusion HD Cloning Kit. CAT with 50 bp homology to pIX ORF upstream and downstream and flanked with SbfI sites. The asterisk represents the site for PacI& EcoRI digestion, blunting, and self-ligation.
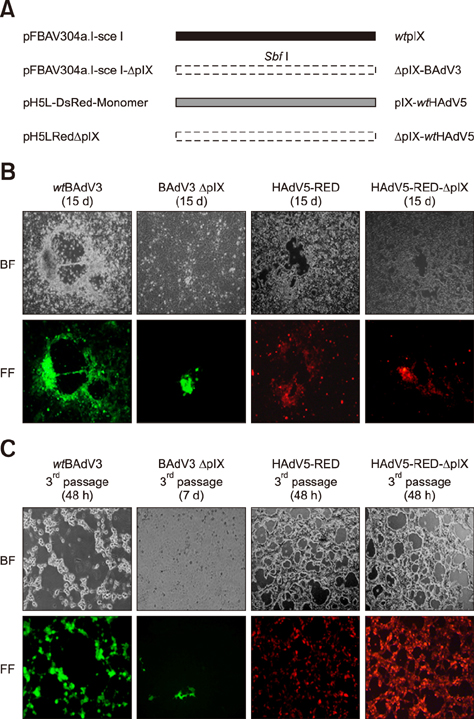
Fig. 2 Generation of pIX-deleted Adenoviruses variants. (A) Schematic representation of the pIX mutants. The pIX-deleted mutants were introduced by homology recombination (pFBAV304a.IsceI-ΔpIX, replace pIX by SbfI enzyme site) and enzyme cleavage (pH5LRedΔpIX, deletion pIX by AflII & MunI digestion and blunt ligation). The full name of genome plasmids are listed on the left and the corresponding pIX mutant proteins expressed by the recombinant viruses were listed on the right. “■” represents the CDS of BAdV3's pIX. “” represents the CDS of HAdV5's pIX, and “□” represents the deletion of the pIX's CDS of BAdV3 or HAdV5. “SbfI” indicates the use of SbfI enzyme site to replace the pIX's CDS of BAdV3. (B) Recombinant virions were rescued either in VIDO DT1 cells (BAdV3) or 293A cells (HAdV5). The cytopathic effect and fluorescence were observed at 15 days post-transfection. (C) The cells (VIDO DT1 or 293A) were infected by 2nd passage of wtBAdV3 or BAdV3ΔpIX virions, or by 2nd passage of HAdV5-RED or HAdV5-RED-ΔpIX virions. BF, bright field; FF, fluorescence field; wt, wild-type.
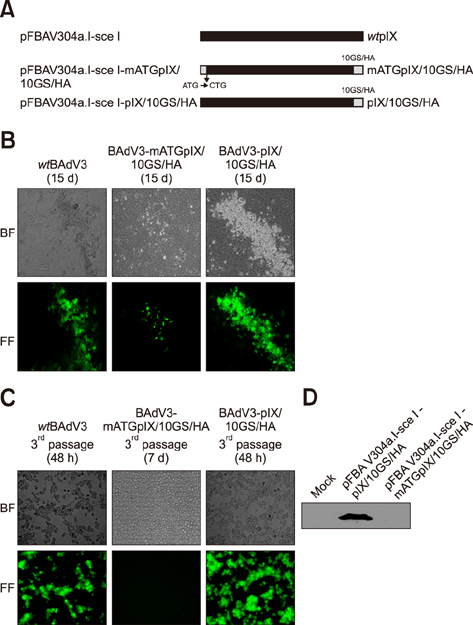
Fig. 3 Generation of start codon mutant and pIX/10GS/HA fusion gene. (A) Schematic representation of the pIX variants. Plasmid names are listed on the left and the corresponding pIX mutant proteins expressed by recombinant viruses are listed on the right. “■” represents the CDS of BAdV3's pIX, “” represents the 10GS/HA linker. “ATG→CTG” indicates the use of CTG to replace the start codon ATG of the BAdV3 pIX. (B) Generation of BAdV3 mutants. Recombinant adenoviral genomes in panel A were transfected into VIDO DT1 cells. At 15 days posttransfection, the cytopathic effect (CPE) and enhanced yellow fluorescent protein (EYFP) were examined. (C) The VIDO DT1 cells were infected by 2nd passage of wtBAdV3, BAdV3- mATGpIX or BAdV3-pIX/10GS/HA virions and observed CPE and EYFP expression. (D) Analysis of the expression of ATG mutant pIXs. The recombination plasmids pFBAV304a.I-sceIpIX/10GS/HA and pFBAV304a.I-sceI-mATGpIX/10GS/HA were transiently transfected in 293T cells and the cell extracts were detected with anti-HA mAb. Mock was a 293T cell extract. BF, bright field; FF, fluorescence field; wt, wild-type.
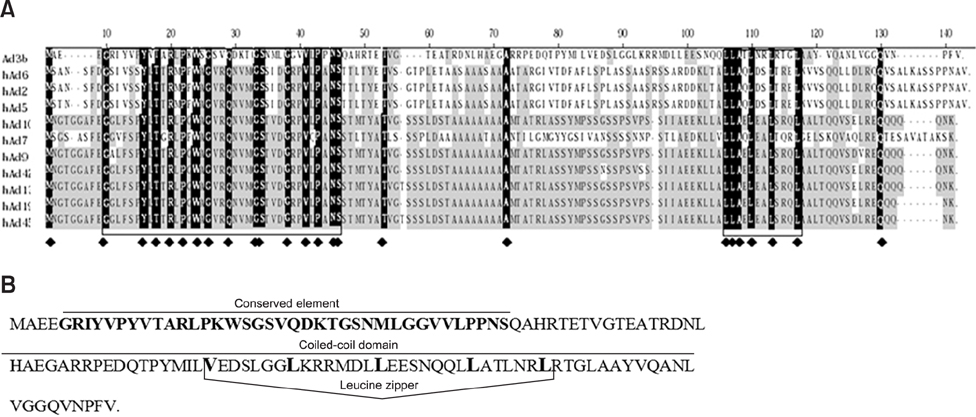
Fig. 4 Conserved amino acid regions in pIX. (A) Amino acid sequence alignment of pIX from Bovine (b) AdV and several human (h) AdV serotypes (Clustal W). Fully conserved residues are shaded black. Residues that occur in more than 50% of the sequences are shaded gray. Conserved sequence elements are boxed. The symbol “♦” denotes identical amino acid residues. (B) Amino acid sequence of BAdV3 pIX. The conserved sequence element and the coiled-coil domain are underlined, and the square bracket delineates the putative leucine zipper (2-ZIP server) [26,27].

Fig. 5 Generation of pIX truncated BAdV3s (BAdV3-pIXΔ1, BAdV3-pIXΔ2, and BAdV3-pIXΔ3). (A) Sequence of the BAdV3 pIX gene. (B) The schematics of truncated pIX mutants. The amino acid positions above the polyline depict the truncated region of pIX. Deletions of the three conserved domains were introduced by using the In-Fusion HD Cloning Kit. The names of genome plasmids are listed on the left and the corresponding names of the pIX mutant proteins expressed by rescued viruses are listed on the right. (C) The genome plasmids in panel B were transfected into VIDO DT1 cells for viral genome rescue, respectively. At 15 days post-transfection, cytopathic effect (CPE) and enhanced yellow fluorescent protein (EYFP) expression were examined. (D) The VIDO DT1 cells were infected by second passage of wtBAdV3, BAdV3-pIXΔ1, BAdV3-pIXΔ2, or BAdV3-pIXΔ3 virions and the CPE and EYFP expression examined. (E) Viral DNAs were extracted from VIDO DT1cells infected with BAdV3-pIXΔ3 or wtBAdV3. After SalI digestion, a 8216 bp fragment in wtBAdV3 viral DNA (lane 2) was cleaved into two fragments (4690 bp and 3550 bp) in BAdV3-pIXΔ3 viral DNA (lane 1), and post-KpnI digestion, a 6730 bp fragment in wtBAdV3 viral DNA (lane 4) was cleaved into two fragments (3578 bp and 3176 bp) in BAdV3-pIXΔ3 viral DNA (lane 3). Marker (M) sizes are shown in bases. (F) Dynamics of wtBAdV3 and BAdV3-pIXΔ3 growth in VIDO DT1 cells. The virus titers were determined at indicated times by plaque-forming unit (PFU) analysis. BF, bright field; FF, fluorescence field; wt, wild-type.
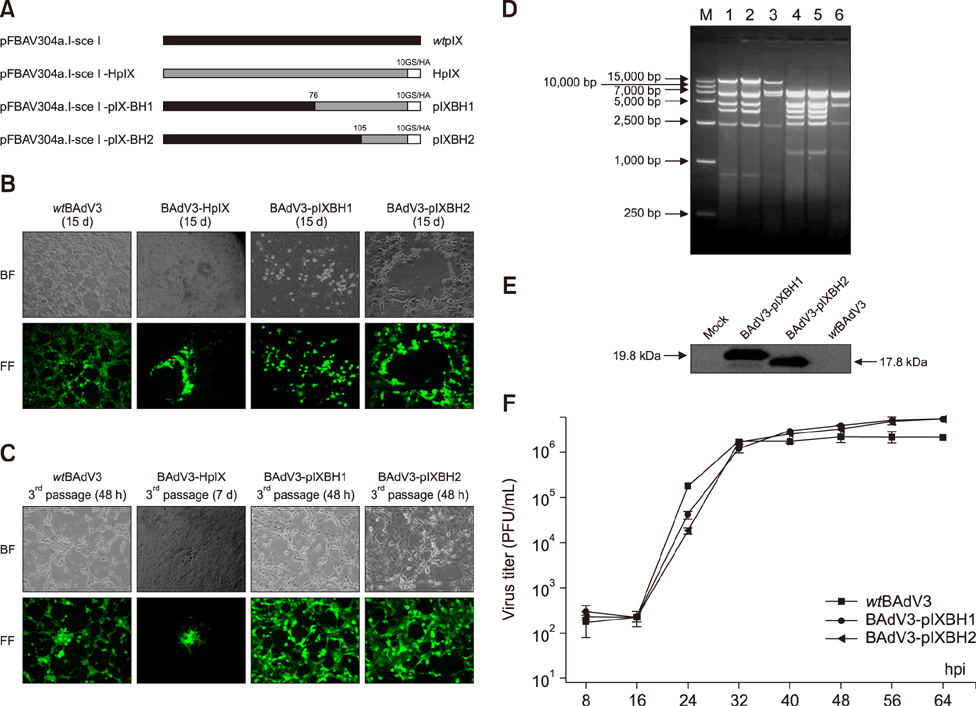
Fig. 6 Generation of chimeric BAdV3s (BAdV3-HpIX, BAdV3-pIXBH1 and BAdV3-pIXBH2). (A) The schematics of chimeric pIXs. “” represents the pIX sequence of BAdV3, “□” represents the pIX sequence of BAdV3 replaced by corresponding region of HAdV5 and “■” represented the 10GS/HA linker. The start amino acid position for chimeric pIX was marked. The names of genome plasmids are listed on the left and the corresponding names of chimeric pIX proteins expressed by rescued viruses are listed on the right. (B) The genome plasmids in panel A were transfected into VIDO DT1 cells for viral genome rescue, respectively. At 15 days post-transfection, the cytopathic effect (CPE) and enhanced yellow fluorescent protein (EYFP) were examined. (C) The VIDO DT1 cells were infected by second passage of wtBAdV3, BAdV3-HpIX, BAdV3-pIXBH1 or BAdV3-pIXBH2 virions and the CPE and EYFP observed. (D) Restriction enzyme analysis of rBAdV3s genome. Viral DNAs were extracted from infected VIDO DT1 cells. After viral DNAs were digested with SalI, a 8216 bp fragment in wtBAdV3 viral DNA (lane 3) was cleaved into two fragments (4700 bp and 3764 bp) in BAdV3-pIXBH1 (lane 1) or two fragments (4694 bp and 3715 bp) in BAdV3-pIXBH2 (lane 2) viral DNA. Post-KpnI digestion, a 6730 bp fragment in wtBAdV3 (lane 6) viral DNA was cleaved into two fragments (3792 bp and 3186 bp) in BAdV3-pIXBH1 (lane 4) or two fragments (3780 bp and 3186 bp ) in BAdV3-pIXBH2 (lane 5) viral DNA. Marker (M) sizes are shown in bases. (E) Western blot analysis of chimeric pIXs. Proteins from uninfected VIDO DT1, BAdV3-pIXBH1-infected VIDO DT1, BAdV3-pIXBH2-infected VIDO DT1, or wtBAdV3-infected VIDO DT1 were separated by SDS-PAGE (15% gel) and transferred to a polyvinylidene fluoride membranes. The separated proteins were probed in western blot by using anti-HA antibody. (F) Dynamics of rBAdV3s growth in VIDO DT1 cells. The virus titers were determined at indicated times by plaque-forming unit (PFU) analysis. BF, bright field; FF, fluorescence field; wt, wild-type.
Reference
-
1. Bangari DS, Shukla S, Mittal SK. Comparative transduction efficiencies of human and nonhuman adenoviral vectors in human, murine, bovine, and porcine cells in culture. Biochem Biophys Res Commun. 2005; 327:960–966.
Article2. Blanchette P, Wimmer P, Dallaire F, Cheng CY, Branton PE. Aggresome formation by the adenoviral protein E1B55K is not conserved among adenovirus species and is not required for efficient degradation of nuclear substrates. J Virol. 2013; 87:4872–4881.
Article3. Cadiñanos J, Bradley A. A Generation of an inducible and optimized piggyBac transposon system. Nucleic Acids Res. 2007; 35:e87.4. Cheng CY, Gilson T, Wimmer P, Schreiner S, Ketner G, Dobner T, Branton PE, Blanchette P. Role of E1B55K in E4orf6/E1B55K E3 ligase complexes formed by different human adenovirus serotypes. J Virol. 2013; 87:6232–6245.
Article5. Colby WW, Shenk T. Adenovirus type 5 virions can be assembled in vivo in the absence of detectable polypeptide IX. J Virol. 1981; 39:977–980.
Article6. Du E, Tikoo SK. Efficient replication and generation of recombinant bovine adenovirus-3 in nonbovine cotton rat lung cells expressing I-SceI endonuclease. J Gene Med. 2010; 12:840–847.
Article7. Fabry CM, Rosa-Calatrava M, Moriscot C, Ruigrok RW, Boulanger P, Schoehn G. The C-terminal domains of adenovirus serotype 5 protein IX assemble into an antiparallel structure on the facets of the capsid. J Virol. 2009; 83:1135–1139.
Article8. Ferreira TB, Alves PM, Aunins JG, Carrondo MJ. Use of adenoviral vectors as veterinary vaccines. Gene Ther. 2005; 12(Suppl 1):S73–S83.
Article9. Furcinitti PS, van Oostrum J, Burnett RM. Adenovirus polypeptide IX revealed as capsid cement by difference images from electron microscopy and crystallography. EMBO J. 1989; 8:3563–3570.
Article10. Ghosh-Choudhury G, Haj-Ahmad Y, Graham FL. Protein IX, a minor component of the human adenovirus capsid, is essential for the packaging of full length genomes. EMBO J. 1987; 6:1733–1739.
Article11. Kunec D, Hanson LA, van Haren S, Nieuwenhuizen IF, Burgess SC. An overlapping bacterial artificial chromosome system that generates vectorless progeny for channel catfish herpesvirus. J Virol. 2008; 82:3872–3881.
Article12. Liu H, Jin L, Koh SBS, Atanasov I, Schein S, Wu L, Zhou ZH. Atomic structure of human adenovirus by cryoEM reveals interactions among protein networks. Science. 2010; 329:1038–1043.
Article13. Lutz P, Rosa-Calatrava M, Kedinger C. The product of the adenovirus intermediate gene IX is a transcriptional activator. J Virol. 1997; 71:5102–5109.
Article14. Parks RJ. Adenovirus protein IX: a new look at an old protein. Mol Ther. 2005; 11:19–25.
Article15. Parks RJ, Graham FL. A helper-dependent system for adenovirus vector production helps define a lower limit for efficient DNA packing. J Virol. 1997; 71:3293–3298.
Article16. Reddy PS, Chen Y, Idamakanti N, Pyne C, Babiuk LA, Tikoo SK. Characterization of early region 1 and pIX of bovine adenovirus-3. Virology. 1999; 253:299–308.
Article17. Reddy PS, Idamakanti N, Zakhartchouk AN, Baxi MK, Lee JB, Pyne C, Babiuk LA, Tikoo SK. Nucleotide sequence, genome organization, and transcription map of bovine adenovirus type 3. J Virol. 1998; 72:1394–1402.
Article18. Rosa-Calatrava M, Grave L, Puvion-Dutilleul F, Chatton B, Kedinger C. Functional analysis of adenovirus protein IX identifies domains involved in capsid stability, transcriptional activity, and nuclear reorganization. J Virol. 2001; 75:7131–7141.
Article19. Rosa-Calatrava M, Puvion-Dutilleul F, Lutz P, Dreyer D, de Thé H, Chatton B, Kedinger C. Adenovirus protein IX sequesters host-cell promyelocytic leukaemia protein and contributes to efficient viral proliferation. EMBO Rep. 2003; 4:969–975.
Article20. Roy A, Kucukural A, Zhang Y. I-TASSER: a unified platform for automated protein structure and function prediction. Nat Protoc. 2010; 5:725–738.
Article21. Sargent KL, Meulenbroek RA, Parks RJ. Activation of adenoviral gene expression by protein IX is not required for efficient virus replication. J Virol. 2004; 78:5032–5037.
Article22. Souquere-Besse S, Pichard E, Filhol O, Legrand V, Rosa-Calatrava M, Hovanessian AG, Cochet C, Puvion-Dutilleul F. Adenovirus infection targets the cellular protein kinase CK2 and RNA-activated protein kinase (PKR) into viral inclusions of the cell nucleus. Microsc Res Tech. 2002; 56:465–478.
Article23. Stewart PL, Fuller SD, Burnett RM. Difference imaging of adenovirus: bridging the resolution gap between X-ray crystallography and electron microscopy. EMBO J. 1993; 12:2589–2599.
Article24. Strunze S, Engelke MF, Wang IH, Puntener D, Boucke K, Schleich S, Way M, Schoenenberger P, Burckhardt CJ, Greber UF. Kinesin-1-mediated capsid disassembly and disruption of the nuclear pore complex promote virus infection. Cell Host Microbe. 2011; 10:210–223.
Article25. van Oostrum J, Burnett RM. Molecular composition of the adenovirus type 2 virion. J Virol. 1985; 56:439–448.
Article26. Vellinga J, van den Wollenberg DJ, van der Heijdt S, Rabelink MJ, Hoeben RC. The coiled-coil domain of the adenovirus type 5 protein IX is dispensable for capsid incorporation and thermostability. J Virol. 2005; 79:3206–3210.
Article27. Zakhartchouk A, Connors W, van Kessel A, Tikoo SK. Bovine adenovirus type 3 containing heterologous protein in the C-terminus of minor capsid protein IX. Virology. 2004; 320:291–300.
Article28. Zakhartchouk AN, Pyne C, Mutwiri GK, Papp Z, Baca-Estrada ME, Griebel P, Babiuk LA, Tikoo SK. Mucosal immunization of calves with recombinant bovine adenovirus-3: induction of protective immunity to bovine herpesvirus-1. J Gen Virol. 1999; 80:1263–1269.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Expression and Packaging of a Human Endogenous Retrovirus-K Genomic DNA Clone
- Construction of Recombinant p53 Adenovirus ( Ad5CMVp53 ) for Cervical Cancer Gene Therapy
- Construction of Recombinant Adeno-Associated Virus Vector (AAVCMVp53) for Human Cervical Cancer Gene Therapy
- Biochemical properties of full-length hepatitis C virus RNA-dependent RNA polymerase expressed in insect cells
- Enhancement of Adenovirus Type 12 Transformation by N-Methyl-N-Nitro-N-Nitrosoguanidine
