In vitro treatment of lipopolysaccharide increases invasion of Pasteurella multocida serotype B:2 into bovine aortic endothelial cells
- Affiliations
-
- 1Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Malaysia. aro@upm.edu.my
- 2Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 Serdang, Malaysia.
- 3Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang, Malaysia.
- KMID: 2407620
- DOI: http://doi.org/10.4142/jvs.2018.19.2.207
Abstract
- Pasteurella multocida serotype B:2 causes hemorrhagic septicemia in cattle and buffalo. The invasion mechanism of the bacterium when invading the bloodstream is unclear. This study aimed to characterize the effects of immunomodulatory molecules, namely dexamethasone and lipopolysaccharide, on the invasion efficiency of P. multocida serotype B:2 toward bovine aortic endothelial cells (BAECs) and the involvement of actin microfilaments in the invasion mechanism. The results imply that treatment of BAECs with lipopolysaccharide at 100 ng/mL for 24 h significantly increases the intracellular bacteria number per cell (p < 0.01) compared with those in untreated and dexamethasone-treated cells. The lipopolysaccharide-treated cells showed a significant decrease in F-actin expression and an increase in G-actin expression (p < 0.001), indicating actin depolymerization of BAECs. However, no significant differences were detected in the invasion efficiency and actin filament reorganization between the dexamethasone-treated and untreated cells. Transmission electron microscopy showed that P. multocida B:2 resided in a vacuolar compartment of dexamethasone-treated and untreated cells, whereas the bacteria resided in cellular membrane of lipopolysaccharide-treated cells. The results suggest that lipopolysaccharide destabilizes the actin filaments of BAECs, which could facilitate the invasion of P. multocida B:2 into BAECs.
Keyword
MeSH Terms
Figure
-
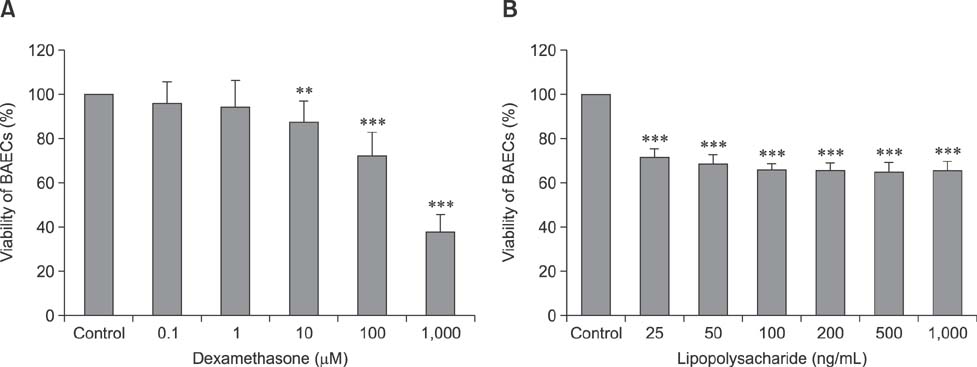
Fig. 1 Effect of dexamethasone (A) and lipopolysaccharide (B) on the viability of bovine aortic endothelial cells (BAECs). The cell viability was assessed by MTT assay. The cells were incubated with the treatments for 24 h. Data are presented as mean ± SD of three independent assays with six replicate samples in each assay. Significant differences compared with untreated cells indicated by **p < 0.01, ***p < 0.001 (Turkey's honest significant difference test, one-way ANOVA).
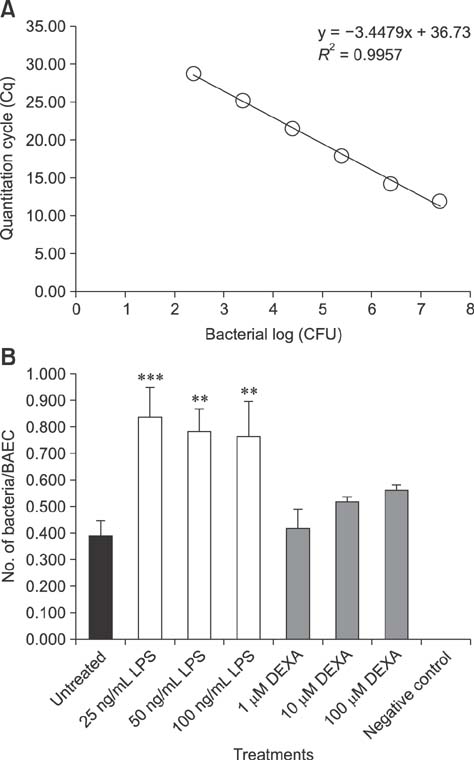
Fig. 2 Quantification of invasion efficiency of bacteria in bovine aortic endothelial cells using real-time polymerase chain reaction assay. The cells were incubated with or without treatments for 24 h and then incubated with bacteria at multiplicity of infection of 100:1 for 2 h. The control was untreated cells and the negative control was DNA of bovine aortic endothelial cells (BAECs). (A) A standard curve was constructed via 10-fold serial dilutions of the extracted genomic DNA of the bacteria from 2.7 × 102 to 2.7 × 10−3 ng. (B) Quantification of bacterial number per cell in different treatments of BAECs. Invasion efficiency is expressed as the bacterial number/BAEC. Data are presented as mean ± SD of two independent assays with at least three replicate samples each. Significant difference from untreated cells indicated by **p < 0.01, ***p < 0.001 (Turkey's honest significant difference test, one-way ANOVA). CFU, colony-forming unit; LPS, lipopolysaccharide; DEXA, dexamethasone.
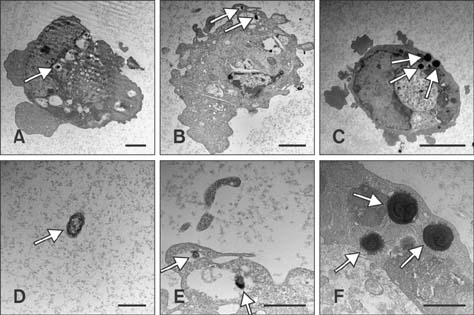
Fig. 3 Micrographs of Pasteurella multocida B:2 infected dexamethasone-, lipopolysaccharide-treated, and untreated bovine aortic endothelial cells (BAECs). The cells were treated with 10 µM dexamethasone (B) or 100 ng/mL lipopolysaccharide (C) for 24 h, and the treated cells were incubated with P. multocida B:2 for 2 h. The untreated cells (A) were used as the control and (D) showed the P. multocida B:2. Micrographs (E) and (F) are enlargements of micrographs (B) and (C), respectively. The white arrows indicate a bacterium. The bacteria resided in vacuolar compartment of dexamethasone-treated and untreated cells, whereas the bacteria resided within the plasma membrane of lipopolysaccharide-treated cells. The samples were observed under transmission electron microscope (H-7100; Hitachi) operating at 100 kV. Scale bars = 1 µm (D-F); 2 µm (A and B); 5 µm (C).
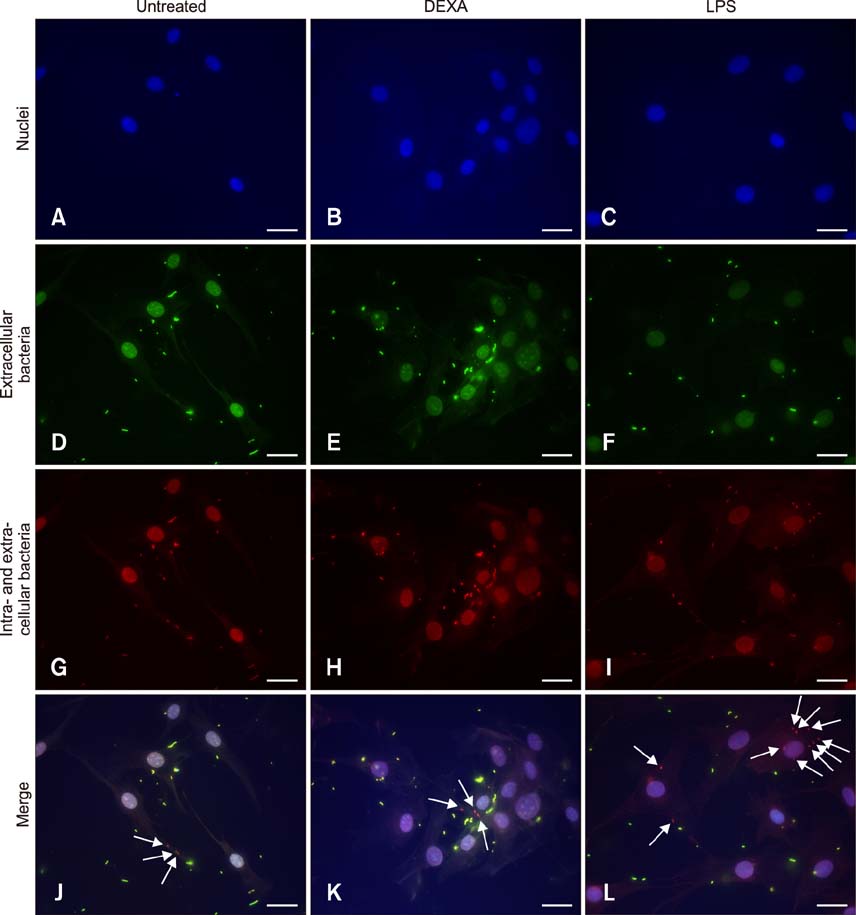
Fig. 4 Images of Pasteurella multocida B:2 infected bovine aortic endothelial cells. The cells were treated with 10 µM DEXA (B, E, H, and K) or 100 ng/mL LPS (C, F, I, and L) for 24 h. The untreated cells (A, D, G, and J) were used as the control. The samples were observed under 60× magnification using an Olympus IX81 microscope. (A-C) Images of cells labeled with DAPI (blue stained nuclei). (D-F) Images of the extracellular bacteria labeled with Alexa Fluor 488 (green stained bacteria). (G-I) Images of intracellular and extracellular bacteria labeled with Alexa Fluor 633 (red stained bacteria). (J-L) Overlapping of the three color images produced images that show the intracellular bacteria (indicated by arrows). Scale bars = 20 µm. DEXA, dexamethasone; LPS, lipopolysaccharide.
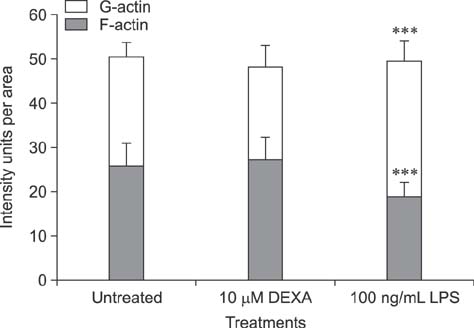
Fig. 5 Effect of dexamethasone (DEXA) and lipopolysaccharide (LPS) on the F- and G-actin pools of bovine aortic endothelial cells. The cells were treated with 10 µM of DEXA or 100 ng/mL of LPS for 24 h. Untreated cells were used as the control. Data are presented as mean ± SD of two independent assays with at least ten replicate samples. Significant differences from untreated cells indicated by ***p < 0.001 (Turkey's honest significant difference test, one-way ANOVA).
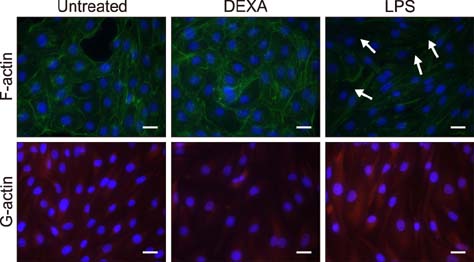
Fig. 6 Images of actin filaments of bovine aortic endothelial cells treated with dexamethasone (DEXA) and lipopolysaccharide (LPS). The cells were incubated with 10 µM DEXA or 100 ng/mL LPS for 24 h. The samples were observed under 60× magnification by using an Olympus IX81 microscope. (A-C) Images of F-actin of the cells labeled with Alexa Fluor 488 Phalloidin (green stained fibers). (D-F) Images of G-actin of the cells labeled with Alexa Fluor 594 DNase I (red fluorescence). All cells were labeled with DAPI (blue stained nuclei). Arrows show loss of the filamentous network of F-actin. Scale bars = 20 µm.
Reference
-
1. Al-haj Ali H, Sawada T, Hatakeyama H, Katayama Y, Ohtsuki N, Itoh O. Invasion of chicken embryo fibroblast cells by avian Pasteurella multocida. Vet Microbiol. 2004; 104:55–62.
Article2. Anderson BH, Watson DL, Colditz IG. The effect of dexamethasone on some immunological parameters in cattle. Vet Res Commun. 1999; 23:399–413.3. Annas S, Zamri-Saad M, Jesse FF, Zunita Z. New sites of localisation of Pasteurella multocida B:2 in buffalo surviving experimental haemorrhagic septicaemia. BMC Vet Res. 2014; 10:88.4. Benkirane A, De Alwis MCL. Haemorrhagic septicaemia, its significance, prevention and control in Asia. Vet Med Czech. 2002; 47:234–240.
Article5. Boyce JD, Adler B. How does Pasteurella multocida respond to the host environment? Curr Opin Microbiol. 2006; 9:117–122.6. Chakravortty D, Koide N, Kato Y, Sugiyama T, Kawai M, Fukada M, Yoshida T, Yokochi T. Cytoskeletal alterations in lipopolysaccharide-induced bovine vascular endothelial cell injury and its prevention by sodium arsenite. Clin Diagn Lab Immunol. 2000; 7:218–225.
Article7. Chen WL, Lin CT, Yao CC, Huang YH, Chou YB, Yin HS, Hu FR. In-vitro effects of dexamethasone on cellular proliferation, apoptosis, and Na+-K+-ATPase activity of bovine corneal endothelial cells. Ocul Immunol Inflamm. 2006; 14:215–223.
Article8. Chung ELT, Abdullah FFJ, Ibrahim HH, Marza AD, Zamri-Saad M, Haron AW, Lila MAM, Norsidin MJ. Clinico-pathology, hematology and biochemistry responses in buffaloes towards Pasteurella multocida type B: 2 immunogen lypopolysaccharide via oral and intravenous routes of infection. Microb Pathog. 2016; 91:141–154.
Article9. De Alwis MCL. Haemorrhagic Septicaemia. ACIAR Monograph No. 57. Canberra: Australian Centre for International Agriculture Research (ACIAR);1999.10. de Alwis MCL, Wijewardana TG, Gomis AIU, Vipulasiri AA. Persistence of the carrier status in haemorrhagic septicaemia (Pasteurella multocida serotype 6:B infection) in buffaloes. Trop Anim Health Prod. 1990; 22:185–194.
Article11. Frey EA, Finlay BB. Lipopolysaccharide induces apoptosis in a bovine endothelial cell line via a soluble CD14 dependent pathway. Microb Pathog. 1998; 24:101–109.
Article12. Galdiero M, De Martino L, Pagnini U, Pisciotta MG, Galdiero E. Interactions between bovine endothelial cells and Pasteurella multocida: association and invasion. Res Microbiol. 2001; 152:57–65.
Article13. Grandel U, Grimminger F. Endothelial responses to bacterial toxins in sepsis. Crit Rev Immunol. 2003; 23:267–299.
Article14. Hardy MP, Gao HB, Dong Q, Ge R, Wang Q, Chai WR, Feng X, Sottas C. Stress hormone and male reproductive function. Cell Tissue Res. 2005; 322:147–153.
Article15. Harper M, Boyce JD, Cox AD, St Michael F, Wilkie IW, Blackall PJ, Adler B. Pasteurella multocida expresses two lipopolysaccharide glycoforms simultaneously, but only a single form is required for virulence: identification of two acceptor-specific heptosyl I transferases. Infect Immun. 2007; 75:3885–3893.
Article16. Horadagoda NU, Hodgson JC, Moon GM, Wijewardana TG, Eckersall PD. Development of a clinical syndrome resembling haemorrhagic septicaemia in the buffalo following intravenous inoculation of Pasteurella multocida serotype B:2 endotoxin and the role of tumour necrosis factor-alpha. Res Vet Sci. 2002; 72:194–200.
Article17. Iovane G, Pagnini P, Galdiero M, Cipollaro de l'Ero G, Vitiello M, D'Isanto M, Marcatili A. Role of Pasteurella multocida porin on cytokine expression and release by murine splenocytes. Vet Immunol Immunopathol. 1998; 66:391–404.
Article18. Jesse FFA, Osman AY, Adamu L, Yusof MSM, Omar AR, Saharee AA, Haron AW, Abdullah R, Zamri-Saad M. Polymerase chain reaction detection of Pasteurella multocida type B:2 in mice following oral inoculation. Asian J Anim Vet Adv. 2013; 8:493–501.19. Kyoui D, Takahashi H, Miya S, Kuda T, Kimura B. Comparison of the major virulence-related genes of Listeria monocytogenes in internalin A truncated strain 36-25-1 and a clinical wild-type strain. BMC Microbiol. 2014; 14:15.
Article20. Lee EJ, Pontes MH, Groisman EA. A bacterial virulence protein promotes pathogenicity by inhibiting the bacterium's own F1Fo ATP synthase. Cell. 2013; 154:146–156.
Article21. Lee MD, Wooley RE, Glisson JR. Invasion of epithelial cell monolayers by turkey strains of Pasteurella multocida. Avian Dis. 1994; 38:72–77.
Article22. Macé S, Mamlouk K, Chipchakova S, Prévost H, Joffraud JJ, Dalgaard P, Pilet MF, Dousset X. Development of a rapid real-time PCR method as a tool to quantify viable Photobacterium phosphoreum bacteria in salmon (Salmo salar) steaks. Appl Environ Microbiol. 2013; 79:2612–2619.
Article23. Mooney A, Byrne C, Clyne M, Johnson-Henry K, Sherman P, Bourke B. Invasion of human epithelial cells by Campylobacter upsaliensis. Cell Microbiol. 2003; 5:835–847.24. Nadkarni MA, Martin FE, Jacques NA, Hunter N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology. 2002; 148:257–266.
Article25. Othman S, Parton R, Coote J. Interaction between mammalian cells and Pasteurella multocida B:2. Adherence, invasion and intracellular survival. Microb Pathog. 2012; 52:353–358.
Article26. Petersen A, Bisgaard M, Townsend K, Christensen H. MLST typing of Pasteurella multocida associated with haemorrhagic septicaemia and development of a real-time PCR specific for haemorrhagic septicaemia associated isolates. Vet Microbiol. 2014; 170:335–341.
Article27. Rafelski SM, Theriot JA. Bacterial shape and ActA distribution affect initiation of Listeria monocytogenes actin-based motility. Biophys J. 2005; 89:2146–2158.
Article28. Rafidah O, Zamri-Saad M. Effect of dexamethasone on protective efficacy of live gdhA derivative Pasteurella multocida B:2 vaccine. Asian J Anim Vet Adv. 2013; 8:548–554.
Article29. Ramdani , Adler B. Opsonic monoclonal antibodies against lipopolysaccharide (LPS) antigens of Pasteurella multocida and the role of LPS in immunity. Vet Microbiol. 1991; 26:335–347.30. Shivachandra SB, Viswas KN, Kumar AA. A review of hemorrhagic septicemia in cattle and buffalo. Anim Health Res Rev. 2011; 12:67–82.
Article31. West NP, Sansonetti P, Mounier J, Exley RM, Parsot C, Guadagnini S, Prévost MC, Prochnicka-Chalufour A, Delepierre M, Tanguy M, Tang CM. Optimization of virulence functions through glucosylation of Shigella LPS. Science. 2005; 307:1313–1317.
Article32. Wheeler R. Outer membrane proteomics of Pasteurella multocida isolates to identify putative host-specificity determinants. Biosci Horiz. 2009; 2:1–12.
Article33. Yap HY, Ghazali K, Wan Mohamad Nazarie WF, Mat Isa MN, Zakaria Z, Omar AR. Draft genome sequence of Pasteurella multocida subsp. multocida strain PMTB, isolated from a buffalo. Genome Announc. 2013; 1:e00872–e00813.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Pasteurella multocida isolation from pigs with respiratory disease in Korea
- A Case of pasteurella multocida pleural empyema
- Hydrogen Peroxide as an Effective Disinfectant for Pasteurella multocida
- Meningoencephalitis and pneumonia caused by Pasteurella multocida in rabbits
- Immunologic reactivity of a lipopolysaccharide-protein complex of type A Pasteurella multocida in mice
