Requirement of CD4 Help for Induction of CD8 T Cell Response Specific for Virally Derived H60
- Affiliations
-
- 1Department of Biomedical Sciences, Seoul National University College of Medicine, Seoul 110-799, Korea. eycii@snu.ac.kr
- 2Therapeutic Antibody Research Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon 305-806, Korea.
- 3Graduate School of Medicine, Korea University, Seoul 136-705, Korea.
- 4Division of Life and Pharmaceutical Sciences, Ewha Woman's University, Seoul 120-750, Korea.
- KMID: 2168010
- DOI: http://doi.org/10.4110/in.2012.12.3.118
Abstract
- CD40-CD40L-mediated help from CD4 T cells is essential to induce primary CD8 T cell responses specific to the non-inflammatory cell-based antigen H60. In this study, using H60 as a model antigen, we generated recombinant vaccinia viruses (rVVs) expressing the H60 CD8 epitope and investigated whether CD4 help was required to activate the CD8 T cell response specific to the virally expressed H60. The immune response after infection with rVVs expressing H60 was similar to that after immunization with H60 congenic splenocytes, with a peak frequency of H60-specific CD8 T cells detected in the blood on day 10 post-infection. A CD8 T cell response specific for virally derived H60 was not induced in CD4-depleted mice, but was in CD40-deficient mice. These results provide insights into the characterization of the CD8 T cell response specifically for antigens originating from cellular sources compared to viral sources.
Figure
-
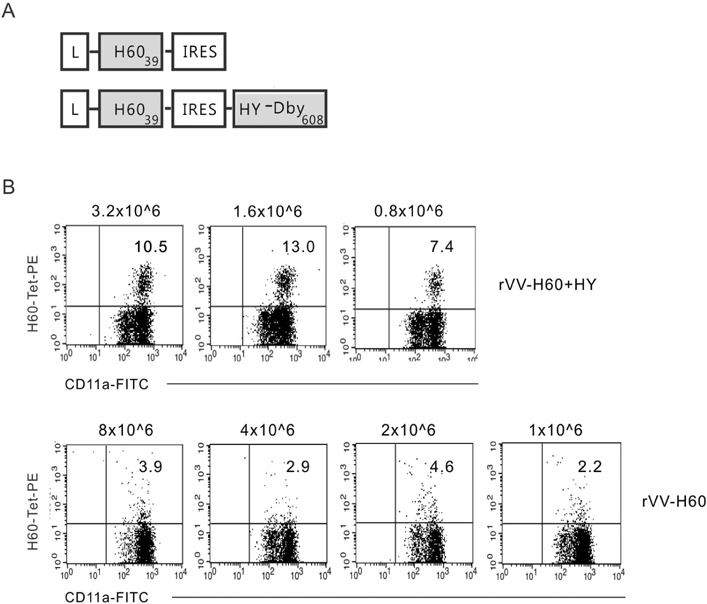
Figure 1 Generation of recombinant vaccinia viruses expressing the H60 CD8 epitope. (A) To generate a recombinant vaccinia virus expressing the H60 CD8 epitope (rVV-H6039), a minigene corresponding to a region encompassing a leader signal peptide to the epitope sequences (39~46 amino acids) of H60 was cloned into a pIRES vector. The DNA fragment encompassing the leader signal to the IRES region containing a multi-cloning site was cloned into a pSC11 vector. rVV-H6039+HY-Dby608 expressed the HY-Dby CD4 epitope peptide (608~622 aa) with an additional Met at the N-terminal, as well as the H60 CD8 epitope. This was achieved by cloning the corresponding minigene into the cloning site located after the IRES region of the rVV-H6039-pSC11 vector. (B) Female B6 mice were immunized with different amounts of rVV-H6039+HY-Dby608 and rVV-H6039 in order to determine the optimal titer of virus needed for the induction of H60-specific CD8 T cell responses, and to compare the ability of each virus to induce the specific immune response. Day 7 post-infection PBLs from immunized mice were stained with H60-Tet-PE, CD11a-FITC, and CD8-APC and analyzed by flow cytometry. Data from the flow cytometry analysis are shown here after gating on CD8+ cells.
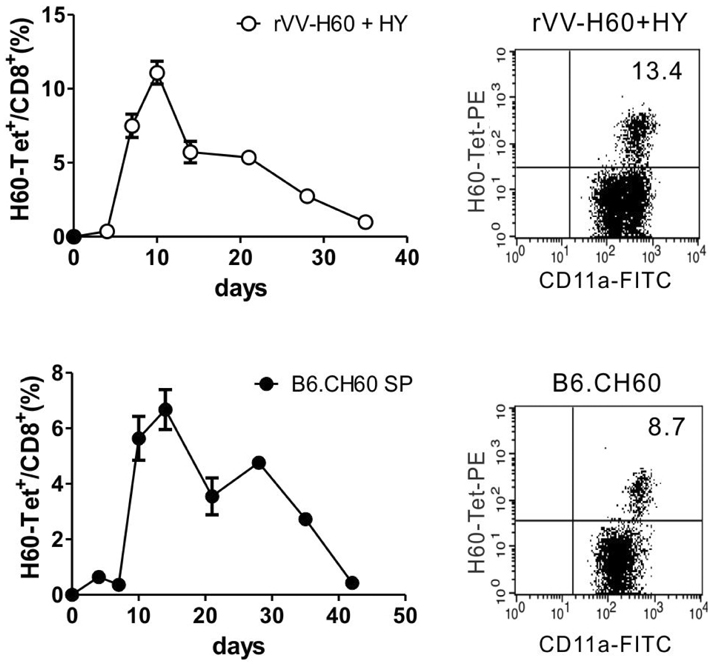
Figure 2 Kinetics of the immune response after infection with rVV-H6039+HY-Dby608. Female B6 mice were immunized with rVV-H6039+HY-Dby608 or male H60 congenic splenocytes and periodic retro-orbital blood samples were obtained. PBLs were stained with H60-Tet-PE, CD11a-FITC, and CD8-APC for flow cytometry. The frequencies of H60-tetramer binding cells in the peripheral blood CD8 T cells at each time point are plotted as kinetics curves. Representative data from flow cytometry performed on day 10 post-immunization are shown after gating on CD8+ cells. These data were obtained from two independent experiments.
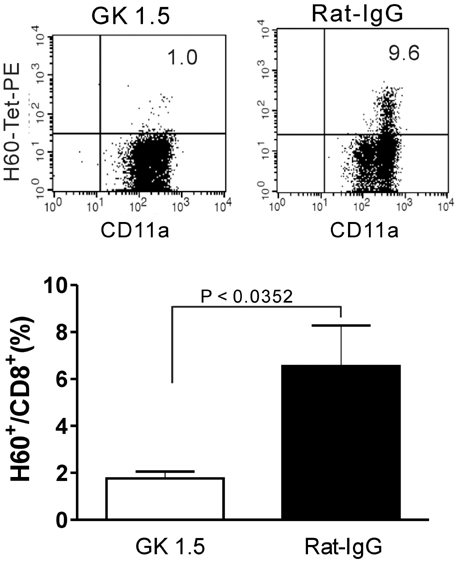
Figure 3 Dependency of the CD8 T cell response to a virally expressed H60 epitope on CD4 help. Female B6 mice were treated with GK1.5 (anti-CD4 mAb) or Rat-IgG (control) three days prior to infection with rVV-H6039+HY-Dby608. PBLs taken periodically from the two different immunized groups were stained with H60-Tet-PE, CD11a-FITC, and CD8-APC. Representative flow cytometry data obtained on day 10 post-infection (peak point) are shown. The peak frequencies observed from four individual mice in each group are plotted. These data were obtained from two independent experiments.

Figure 4 The CD8 T cell response specific for virally expressed H60 is independent of CD40. This figure shows female CD40-deficient and wild type B6 mice immunized with rVV-H6039+HY-Dby608 (A) and female CD40-deficient and wild type B6 mice immunized with male H60 congenic splenocytes (B). PBLs taken periodically from immunized mice were stained with H60-Tet-PE, CD11a-FITC, and CD8-APC. Representative flow cytometry data obtained on day 10 post-infection (peak point) are shown. The peak frequencies observed from five individual mice in each group are plotted. These data were obtained from two independent experiments.
Reference
-
1. Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: implications for vaccine development. Nat Rev Immunol. 2002. 2:251–262.
Article2. Keene JA, Forman J. Helper activity is required for the in vivo generation of cytotoxic T lymphocytes. J Exp Med. 1982. 155:768–782.
Article3. Rees MA, Rosenberg AS, Munitz TI, Singer A. In vivo induction of antigen-specific transplantation tolerance to Qa1a by exposure to alloantigen in the absence of T-cell help. Proc Natl Acad Sci U S A. 1990. 87:2765–2769.4. Ryu SJ, Jung KM, Yoo HS, Kim TW, Kim S, Chang J, Choi EY. Cognate CD4 help is essential for the reactivation and expansion of CD8 memory T cells directed against the hematopoietic cell-specific dominant minor histocompatibility antigen, H60. Blood. 2009. 113:4273–4280.5. Bennett SR, Carbone FR, Karamalis F, Flavell RA, Miller JF, Heath WR. Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature. 1998. 393:478–480.
Article6. Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 1998. 393:480–483.
Article7. Ridge JP, Di Rosa F, Matzinger P. A conditioned dendritic cell can be a temporal bridge between a CD4+ T-helper and a T-killer cell. Nature. 1998. 393:474–478.
Article8. Jung KM, Choi EY. Role for CD40 and CD40L Expression in Generating CD8 T Cell Response to Minor Histcompatibility Antigen, H60. Immune Netw. 2007. 7:173–178.
Article9. Rahemtulla A, Fung-Leung WP, Schilham MW, Kündig TM, Sambhara SR, Narendran A, Arabian A, Wakeham A, Paige CJ, Zinkernagel RM, Miller RG. Normal development and function of CD8+ cells but markedly decreased helper cell activity in mice lacking CD4. Nature. 1991. 353:180–184.
Article10. Bevan MJ. Helping the CD8(+) T-cell response. Nat Rev Immunol. 2004. 4:595–602.
Article11. Janssen EM, Lemmens EE, Wolfe T, Christen U, von Herrath MG, Schoenberger SP. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature. 2003. 421:852–856.
Article12. Shedlock DJ, Shen H. Requirement for CD4 T cell help in generating functional CD8 T cell memory. Science. 2003. 300:337–339.
Article13. Sun JC, Bevan MJ. Defective CD8 T cell memory following acute infection without CD4 T cell help. Science. 2003. 300:339–342.
Article14. Roopenian D, Choi EY, Brown A. The immunogenomics of minor histocompatibility antigens. Immunol Rev. 2002. 190:86–94.
Article15. Choi EY, Yoshimura Y, Christianson GJ, Sproule TJ, Malarkannan S, Shastri N, Joyce S, Roopenian DC. Quantitative analysis of the immune response to mouse non-MHC transplantation antigens in vivo: the H60 histocompatibility antigen dominates over all others. J Immunol. 2001. 166:4370–4379.
Article16. Choi EY, Christianson GJ, Yoshimura Y, Sproule TJ, Jung N, Joyce S, Roopenian DC. Immunodominance of H60 is caused by an abnormally high precursor T cell pool directed against its unique minor histocompatibility antigen peptide. Immunity. 2002. 17:593–603.
Article17. Malarkannan S, Shih PP, Eden PA, Horng T, Zuberi AR, Christianson G, Roopenian D, Shastri N. The molecular and functional characterization of a dominant minor H antigen, H60. J Immunol. 1998. 161:3501–3509.
Article18. Warren EH, Fujii N, Akatsuka Y, Chaney CN, Mito JK, Loeb KR, Gooley TA, Brown ML, Koo KK, Rosinski KV, Ogawa S, Matsubara A, Appelbaum FR, Riddell SR. Therapy of relapsed leukemia after allogeneic hematopoietic cell transplantation with T cells specific for minor histocompatibility antigens. Blood. 2010. 115:3869–3878.
Article19. Choi JH, Ryu SJ, Jung KM, Kim S, Chang J, Kim TW, Choi EY. TCR diversity of H60-specific CD8 T cells during the response evolution and influence of CD4 help. Transplantation. 2009. 87:1609–1616.20. Harrington LE, Most Rv R, Whitton JL, Ahmed R. Recombinant vaccinia virus-induced T-cell immunity: quantitation of the response to the virus vector and the foreign epitope. J Virol. 2002. 76:3329–3337.21. Xu R, Johnson AJ, Liggitt D, Bevan MJ. Cellular and humoral immunity against vaccinia virus infection of mice. J Immunol. 2004. 172:6265–6271.
Article22. Norbury CC, Princiotta MF, Bacik I, Brutkiewicz RR, Wood P, Elliott T, Bennink JR, Yewdell JW. Multiple antigen-specific processing pathways for activating naive CD8+ T cells in vivo. J Immunol. 2001. 166:4355–4362.
Article23. Novy P, Quigley M, Huang X, Yang Y. CD4 T cells are required for CD8 T cell survival during both primary and memory recall responses. J Immunol. 2007. 179:8243–8251.
Article24. Agnellini P, Wiesel M, Schwarz K, Wolint P, Bachmann MF, Oxenius A. Kinetic and mechanistic requirements for helping CD8 T cells. J Immunol. 2008. 180:1517–1525.
Article25. Bacik I, Cox JH, Anderson R, Yewdell JW, Bennink JR. TAP (transporter associated with antigen processing)-independent presentation of endogenously synthesized peptides is enhanced by endoplasmic reticulum insertion sequences located at the amino- but not carboxyl-terminus of the peptide. J Immunol. 1994. 152:381–387.
Article26. Hwang ML, Lukens JR, Bullock TN. Cognate memory CD4+ T cells generated with dendritic cell priming influence the expansion, trafficking, and differentiation of secondary CD8+ T cells and enhance tumor control. J Immunol. 2007. 179:5829–5838.
Article27. Xiao Z, Curtsinger JM, Prlic M, Jameson SC, Mescher MF. The CD8 T cell response to vaccinia virus exhibits site-dependent heterogeneity of functional responses. Int Immunol. 2007. 19:733–743.
Article28. Moutaftsi M, Bui HH, Peters B, Sidney J, Salek-Ardakani S, Oseroff C, Pasquetto V, Crotty S, Croft M, Lefkowitz EJ, Grey H, Sette A. Vaccinia virus-specific CD4+ T cell responses target a set of antigens largely distinct from those targeted by CD8+ T cell responses. J Immunol. 2007. 178:6814–6820.
Article29. Yang Y, Xiang Z, Ertl HC, Wilson JM. Upregulation of class I major histocompatibility complex antigens by interferon gamma is necessary for T-cell-mediated elimination of recombinant adenovirus-infected hepatocytes in vivo. Proc Natl Acad Sci U S A. 1995. 92:7257–7261.
Article30. Van Deusen KE, Rajapakse R, Bullock TN. CD70 expression by dendritic cells plays a critical role in the immunogenicity of CD40-independent, CD4+ T cell-dependent, licensed CD8+ T cell responses. J Leukoc Biol. 2010. 87:477–485.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Role for CD40 and CD40L Expression in Generating CD8 T Cell Response to Minor Histcompatibility Antigen, H60
- Subdominant H60 antigen-specific CD8 T-cell response precedes dominant H4 antigen-specific response during the initial phase of allogenic skin graft rejection
- The Role of CD4 T Cell Help in CD8 T Cell Differentiation and Function During Chronic Infection and Cancer
- Selection of Thymocytes Expressing Transgenic TCR Specific for a Minor Histocompatibility Antigen, H60
- The effect of interleukin 2 on the induction Of Nk 1.1 expression in CD8+ and CD4-CD8-T Cell
