Extracellular Vimentin Alters Energy Metabolism And Induces Adipocyte Hypertrophy
- Affiliations
-
- 1Department of Medicine, Graduate School, Ewha Womans University, Seoul, Korea
- 2Department of Molecular Medicine, Ewha Womans University College of Medicine, Seoul, Korea
- KMID: 2553593
- DOI: http://doi.org/10.4093/dmj.2022.0332
Abstract
- Background
Previous studies have reported that oxidative stress contributes to obesity characterized by adipocyte hypertrophy. However, mechanism has not been studied extensively. In the current study, we evaluated role of extracellular vimentin secreted by oxidized low-density lipoprotein (oxLDL) in energy metabolism in adipocytes.
Methods
We treated 3T3-L1-derived adipocytes with oxLDL and measured vimentin which was secreted in the media. We evaluated changes in uptake of glucose and free fatty acid, expression of molecules functioning in energy metabolism, synthesis of adenosine triphosphate (ATP) and lactate, markers for endoplasmic reticulum (ER) stress and autophagy in adipocytes treated with recombinant vimentin.
Results
Adipocytes secreted vimentin in response to oxLDL. Microscopic evaluation revealed that vimentin treatment induced increase in adipocyte size and increase in sizes of intracellular lipid droplets with increased intracellular triglyceride. Adipocytes treated with vimentin showed increased uptake of glucose and free fatty acid with increased expression of plasma membrane glucose transporter type 1 (GLUT1), GLUT4, and CD36. Vimentin treatment increased transcription of GLUT1 and hypoxia-inducible factor 1α (Hif-1α) but decreased GLUT4 transcription. Adipose triglyceride lipase (ATGL), peroxisome proliferator-activated receptor γ (PPARγ), sterol regulatory element-binding protein 1 (SREBP1), diacylglycerol O-acyltransferase 1 (DGAT1) and 2 were decreased by vimentin treatment. Markers for ER stress were increased and autophagy was impaired in vimentin-treated adipocytes. No change was observed in synthesis of ATP and lactate in the adipocytes treated with vimentin.
Conclusion
We concluded that extracellular vimentin regulates expression of molecules in energy metabolism and promotes adipocyte hypertrophy. Our results show that vimentin functions in the interplay between oxidative stress and metabolism, suggesting a mechanism by which adipocyte hypertrophy is induced in oxidative stress.
Keyword
Figure
-

Fig. 1. Oxidized low-density lipoprotein (oxLDL) induces vimentin secretion in 3T3-L1-derived adipocytes. (A) 3T3-L1-derived adipocytes were treated with or without oxLDL (50 µg/mL) for 8, 16, 24 hours and the concentrations of extracellular vimentin in the media were measured using enzyme-linked immunosorbent assay (ELISA). Data were normalized to the protein amount (mg) of the adipocytes used in the assay (n=3). (B) Detection of vimentin in the concentrated media of 3T3-L1-adipocytes cultured with LDL (50 µg/mL) or different concentrations of oxLDL (25, 50 µg/mL) for 24 hours. In the Western blot, His (molecular weight, 1.6 kDa)-tagged recombinant vimentin was used for a positive control. Data were normalized to the protein amount (mg) of the adipocytes obtained from the individual culture dish (n=3). (C) Detection of vimentin in the concentrated media of 3T3-L1-adipocytes cultured with LDL (50 µg/mL for 24 hours) or oxLDL (50 µg/mL for 8, 16, and 24 hours) using Western blotting. Data were normalized to the cellular protein (mg) (n=3). (D) 3T3-L1-adipocytes were pretreated with anti-CD36 antibody (2 µg/mL) before incubating with LDL (50 µg/mL for 24 hours) or oxLDL (50 µg/mL for 24 hours). Detection of vimentin in the concentrated media of the adipocytes using Western blotting and data were normalized to the cellular protein (mg) (n=3). (E) 3T3-L1-adipocytes were pretreated with or without H-89 (10 µM) before incubating with LDL (50 µg/mL for 24 hours) or oxLDL (50 µg/mL for 24 hours). Detection of vimentin in the concentrated media of adipocytes using Western blotting and data were normalized to the cellular protein (mg) (n=3). All experiments were done more than three times. One-way analysis of variance (ANOVA) with Bonferroni post hoc test was done. SF, serum free. aP<0.05, bP<0.01, cP<0.001.
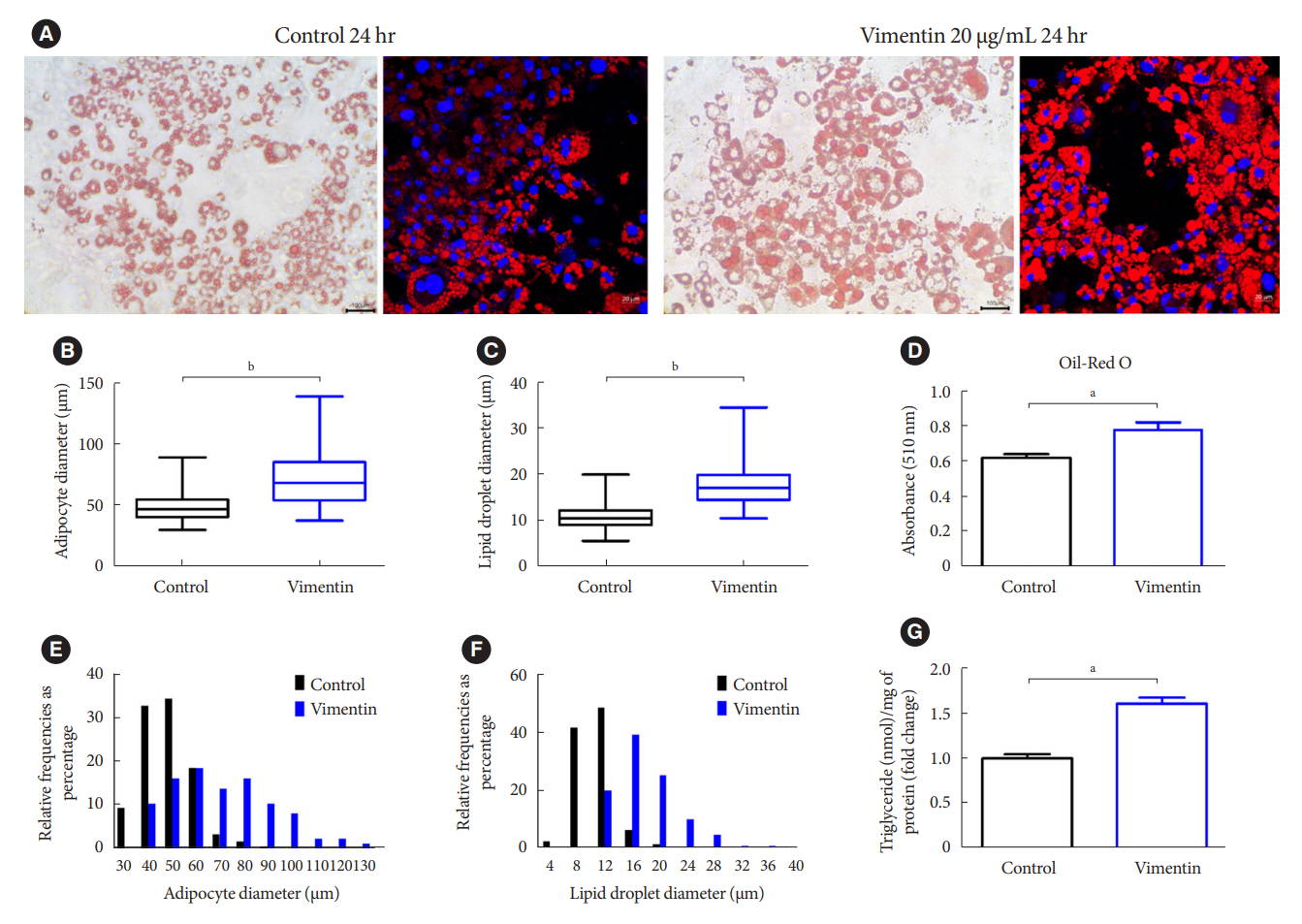
Fig. 2. Extracellular vimentin induces adipocyte hypertrophy. (A) 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours, were stained with Oil-Red O and 4ʹ,6-diamidino-2-phenylindole (DAPI). Light microscopy (left panel) and fluorescence microscopy (right panel). Scale bar: 100 µm (left panel), 20 µm (right panel). (B) Comparison of adipocyte diameter (µm) between vimentin-treated (vimentin) and untreated (control) adipocytes. (C) Comparison of lipid droplet diameter (µm) between vimentin-treated (vimentin) and untreated (control) adipocytes. (D) Absorbance of the Oil-Red O dye extracted from the 3T3-L1-derived adipocytes in (A) (n=3). (E) Size distribution of 3T3-L1 adipocytes treated with or without vimentin (20 µg/mL) for 24 hours. (F) Size distribution of intracellular lipid droplets in the 3T3-L1 adipocytes treated with or without vimentin (20 µg/mL) for 24 hours. (G) Intracellular triglyceride was measured using 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Data were normalized to the amount (mg) of total protein from the adipocytes used in the assay (n=3). aP<0.05, bP<0.001.
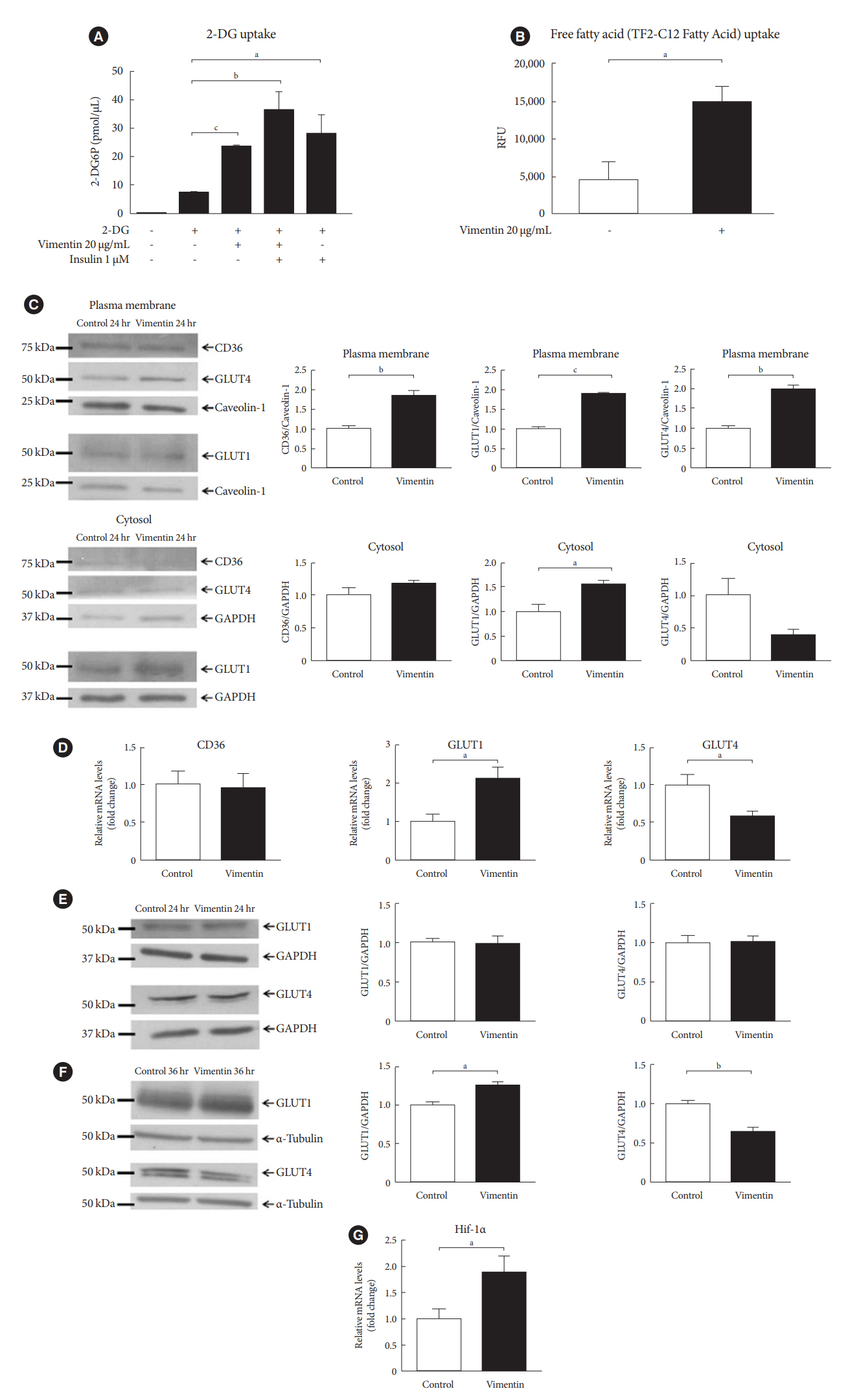
Fig. 3. Extracellular vimentin promotes glucose uptake and free fatty acid (FFA) uptake via changes in expression of glucose transporters and fatty acid transporter. (A) 3T3-L1-derived adipocytes were treated with recombinant vimentin (20 µg/mL for 24 hours), insulin (1 µM for 30 minutes) or both vimentin (20 µg/mL for 24 hours) and insulin (1 µM for 30 minutes). 2-Deoxyglucose (2-DG) uptake was measured (n=4). (B) 3T3-L1-derived adipocytes were treated with or without recombinant vimentin (20 µg/mL) for 24 hours and FFA uptake was measured using fluorometric fatty acid dye-loading solution (TF2-C12 Fatty Acid) uptake assay (n=3). (C) Western blot analyses for CD36, glucose transporter type 4 (GLUT4), and GLUT1 were performed using fractionated lysates (plasma membrane and cytosol) of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Band quantification was performed using either caveolin-1 (BD Biosciences) as a plasma membrane marker, or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a cytosol marker (n=3). (D) Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) analyses for CD36, GLUT1, and GLUT4 were performed using RNA from 3T3-L1-derived adipocytes cultured with or without recombinant vimentin (20 µg/mL) for 24 hours. GAPDH was used as an internal control (n=5, n=6, n=4). (E) Western blot analyses for GLUT1 and GLUT4 using total lysates of 3T-3L1-derived adipocytes treated with or with recombinant vimentin (20 µg/mL) for 24 hours. Band quantification was performed using GAPDH, as an internal control (n=3). (F) Western blot analyses for GLUT1 and GLUT4 were performed using total lysates of 3T3-L1-derived adipocytes with or with recombinant vimentin (20 µg/mL) for 36 hours. Band quantification was performed using α-tubulin (AbFrontier), an internal control (n=3). (G) qRT-PCR analysis for hypoxia-inducible factor 1α (Hif-1α) was performed using RNA from 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. GAPDH was used as an internal control (n=4). RFU, relative fluorescence unit. aP<0.05, bP<0.01, cP<0.001.
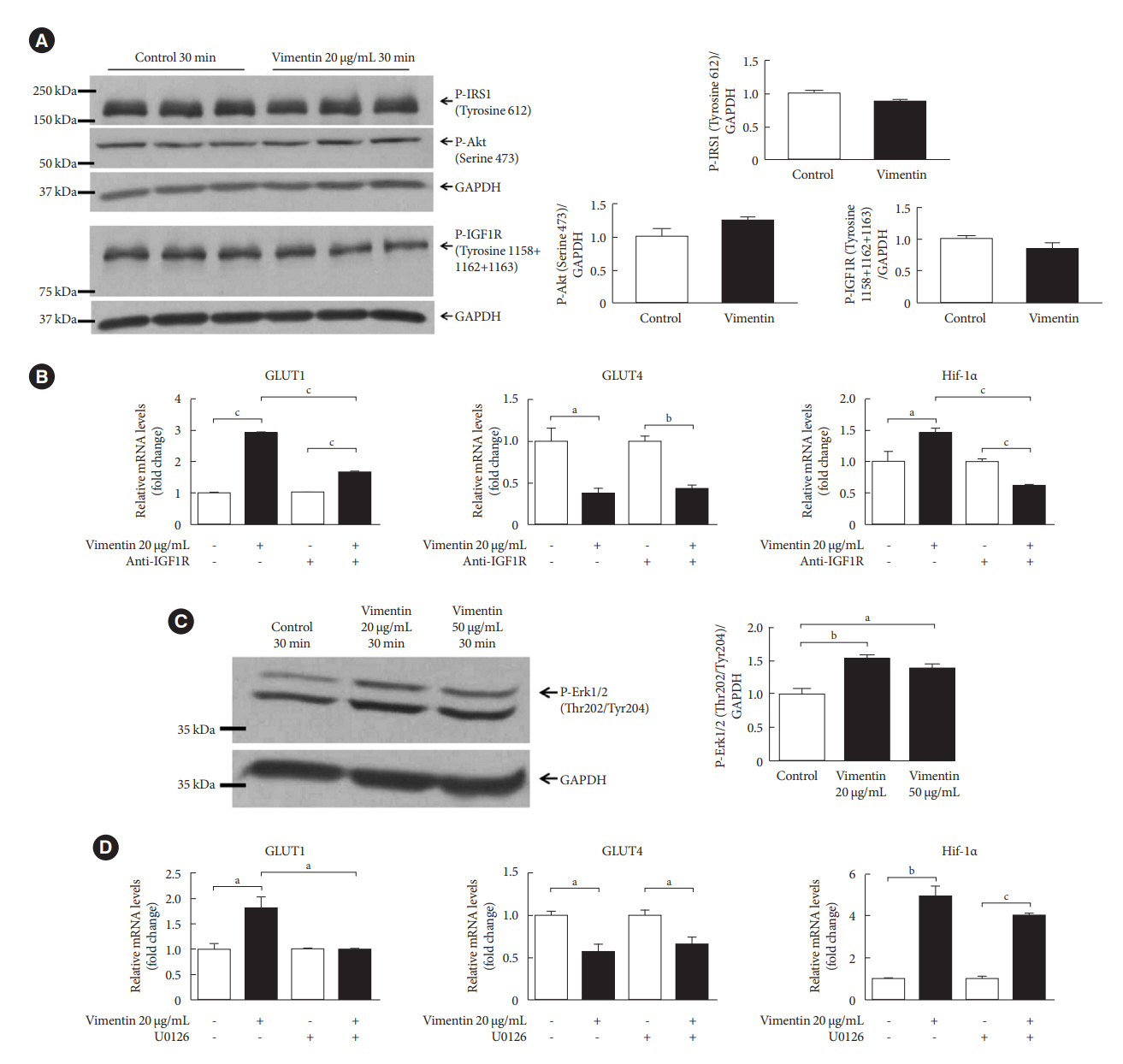
Fig. 4. Extracellular vimentin-induced glucose transporter type 1 (GLUT1) expression depends on insulin-like growth factor 1 receptor (IGF1R) and activation of extracellular-signal-regulated kinase (ERK). (A) Western blot analyses for phospho-insulin receptor substrate 1 (P-IRS1; Tyrosine 612), P-Akt (Serine 473), and P-IGF1R (Tyrosine 1158+1162+1163) were performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 30 minutes. Band quantification was performed using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal control. Samples from three different batches of treatment were loaded onto a gel (n=3). (B) Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) analyses for GLUT1, GLUT4, hypoxia-inducible factor 1α (Hif-1α) were performed with RNA from 3T3-L1-derived adipocytes treated or untreated with anti-IGF1R antibody (11 µg/mL) for 1 hour and then incubated with or without recombinant vimentin (20 µg/mL) for 24 hours. GAPDH was used as an internal control (GLUT1, n=3; GLUT4, n=3; Hif-1α, n=4). (C) Western blotting for P-ERK1/2 (Threonine 202/Tyrosine 204) was performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 30 minutes. Band quantification was performed using GAPDH as an internal control (n=3). (D) qRT-PCR analyses for GLUT1, GLUT4, Hif-1α were performed with RNA from 3T3-L1-derived adipocytes treated or untreated with U0126 (ERK inhibitor, 25 µM) for 1 hour and then incubated with or without recombinant vimentin (20 µg/mL) for 24 hours. GAPDH was used as an internal control (n=3 for each). aP<0.05, bP<0.01, cP<0.001.
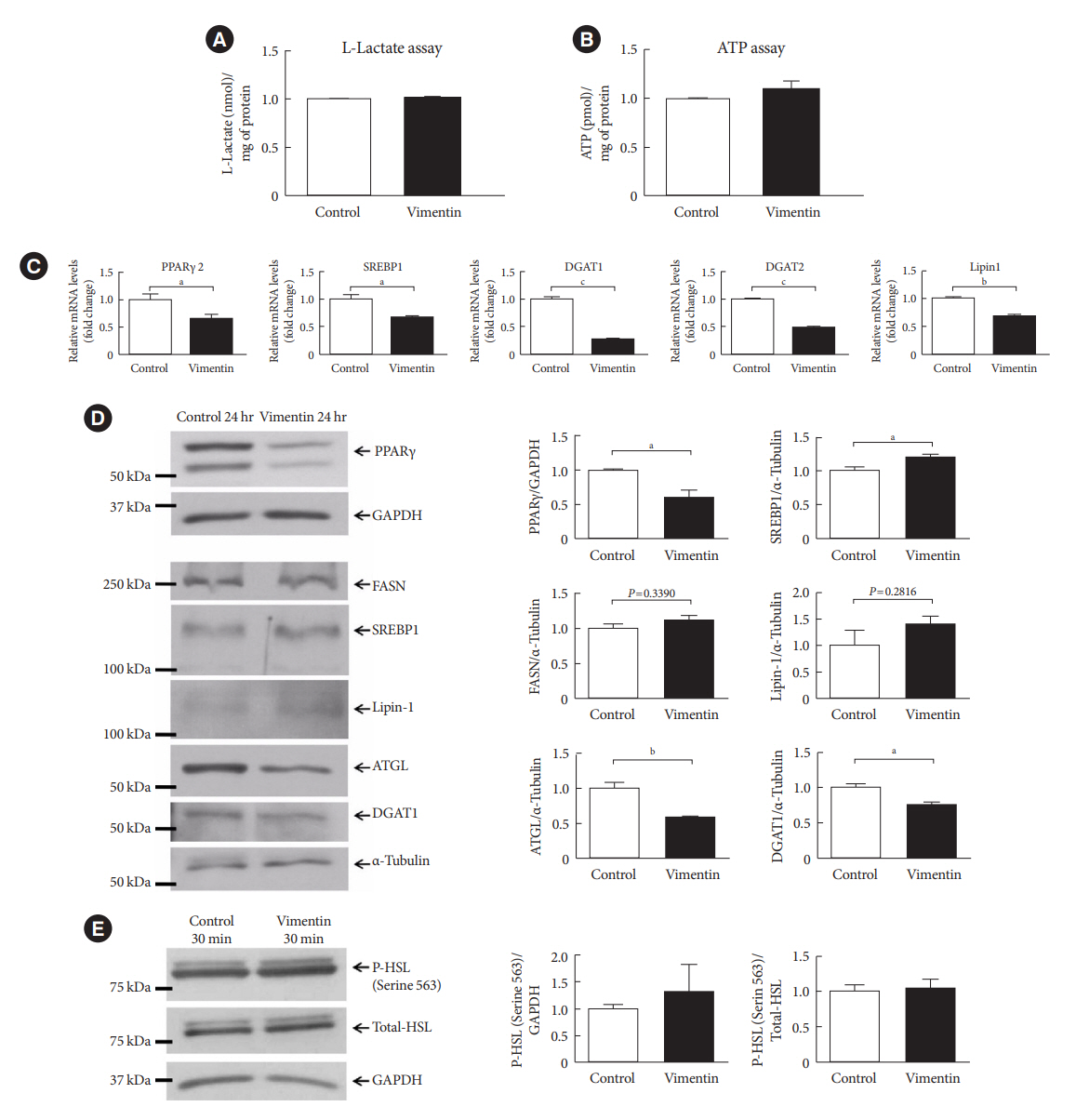
Fig. 5. Extracellular vimentin modulates expression of molecules for lipogenesis and lipolysis. (A) L-Lactate was measured in the culture media of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Data were normalized to the protein amount (mg) of the adipocytes used in the assay (n=2). (B) Intracellular adenosine triphosphate (ATP) was measured in 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Data were normalized to the protein amount (mg) of the adipocytes used in the assay (n=4). (C) Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) analyses for peroxisome proliferator-activated receptor γ 2 (PPARγ2), sterol regulatory element-binding protein 1 (SREBP1), diacylglycerol O-acyltransferase 1 (DGAT1), DGAT2, and Lipin1 were performed using RNA from 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control (n=4, n=3, n=3, n=3). (D) Western blot analysis for PPARγ, fatty acid synthase (FASN), SREBP1, Lipin1, adipose triglyceride lipase (ATGL), and DGAT1 were performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Band quantification was performed using GAPDH and α-tubulin as internal controls (n=3). (E) Western blot analysis for phosphor-hormone-sensitive lipase (P-HSL; Serine 564) was performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 30 minutes. Band quantification was performed using total HSL and GAPDH as internal controls (n=3). aP<0.05, bP<0.01, cP<0.001.
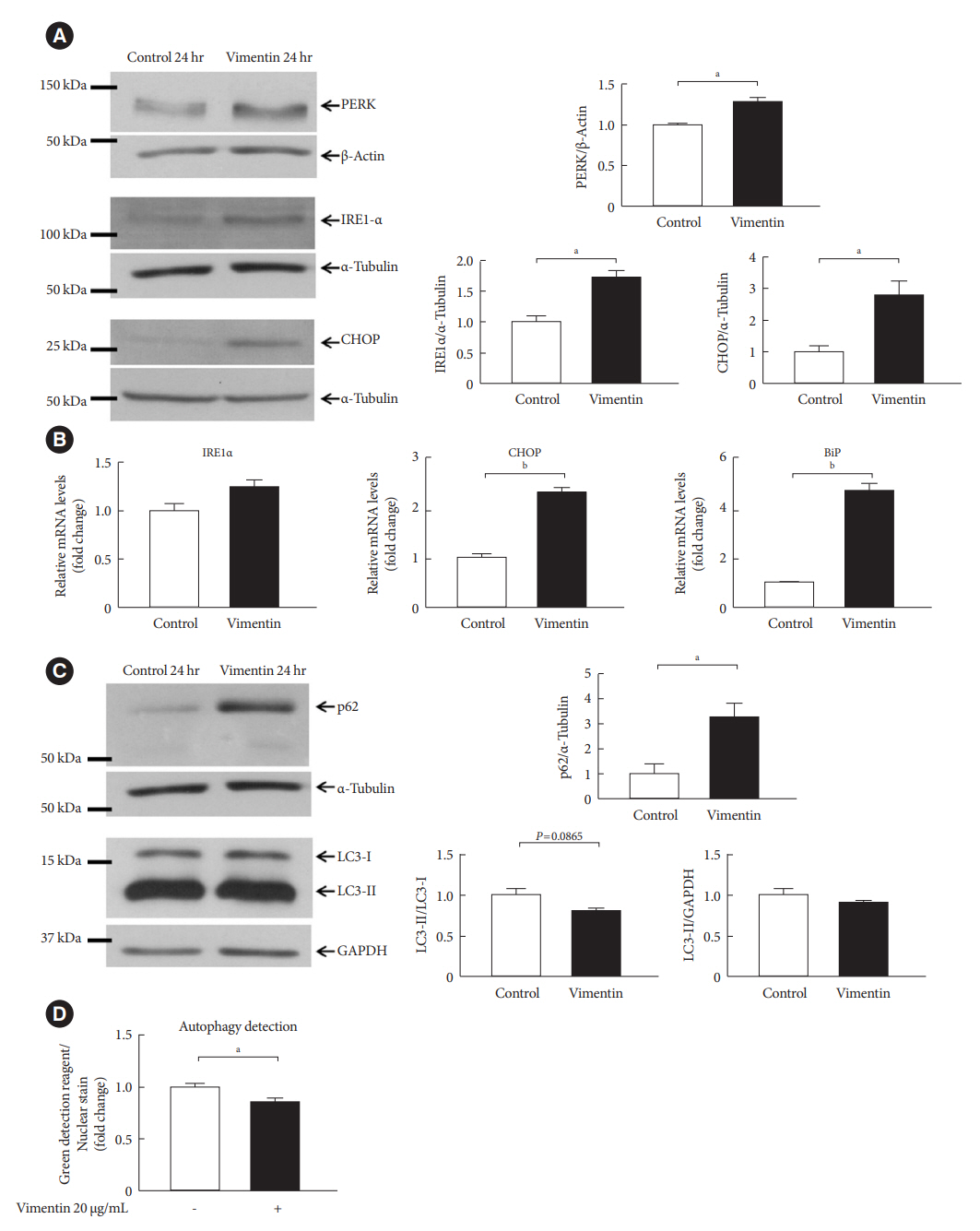
Fig. 6. Extracellular vimentin induces endoplasmic reticulum (ER) stress and impairs autophagy. (A) Western blot analyses for protein kinase R-like ER kinase (PERK), inositol-requiring enzyme 1 α (IRE1α), and C/EBP homologous protein (CHOP) were performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Band quantification was performed using β-actin and α-tubulin as internal controls (n=3). (B) Quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) analyses for IRE1α, CHOP, and binding immunoglobulin protein (BiP) were performed using RNA from 3T3-L1-derived adipocytes treated with or without vimentin (20 µg/mL) for 24 hours. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control (n=4). (C) Western blot analysis for p62, light chain 3 (LC3)-I, and LC3-II were performed using lysates of 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours. Band quantification was performed using α-tubulin and GAPDH as internal controls (n=3). (D) Autophagic vacuoles in 3T3-L1-derived adipocytes treated with or without recombinant vimentin (20 µg/mL) for 24 hours, were measured. Data were normalized to Hoechst 33342 staining in the adipocytes used in the assay (n=5). aP<0.05, bP<0.001.
Reference
-
1. Grundy SM. Obesity, metabolic syndrome, and cardiovascular disease. J Clin Endocrinol Metab. 2004; 89:2595–600.
Article2. Garcia-Sanchez A, Gamez-Nava JI, Diaz-de la Cruz EN, Cardona-Munoz EG, Becerra-Alvarado IN, Aceves-Aceves JA, et al. The effect of visceral abdominal fat volume on oxidative stress and proinflammatory cytokines in subjects with normal weight, overweight and obesity. Diabetes Metab Syndr Obes. 2020; 13:1077–87.3. Pennathur S, Heinecke JW. Mechanisms of oxidative stress in diabetes: implications for the pathogenesis of vascular disease and antioxidant therapy. Front Biosci. 2004; 9:565–74.
Article4. Lee H, Lee YJ, Choi H, Ko EH, Kim JW. Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion. J Biol Chem. 2009; 284:10601–9.
Article5. Njajou OT, Kanaya AM, Holvoet P, Connelly S, Strotmeyer ES, Harris TB, et al. Association between oxidized LDL, obesity and type 2 diabetes in a population-based cohort, the Health, Aging and Body Composition Study. Diabetes Metab Res Rev. 2009; 25:733–9.
Article6. Coelho M, Oliveira T, Fernandes R. Biochemistry of adipose tissue: an endocrine organ. Arch Med Sci. 2013; 9:191–200.7. Kawai T, Autieri MV, Scalia R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021; 320:C375–91.
Article8. Muir LA, Neeley CK, Meyer KA, Baker NA, Brosius AM, Washabaugh AR, et al. Adipose tissue fibrosis, hypertrophy, and hyperplasia: correlations with diabetes in human obesity. Obesity (Silver Spring). 2016; 24:597–605.
Article9. Heid H, Rickelt S, Zimbelmann R, Winter S, Schumacher H, Dorflinger Y, et al. On the formation of lipid droplets in human adipocytes: the organization of the perilipin-vimentin cortex. PLoS One. 2014; 9:e90386.
Article10. Kim S, Kim I, Cho W, Oh GT, Park YM. Vimentin deficiency prevents high-fat diet-induced obesity and insulin resistance in mice. Diabetes Metab J. 2021; 45:97–108.
Article11. Mor-Vaknin N, Punturieri A, Sitwala K, Markovitz DM. Vimentin is secreted by activated macrophages. Nat Cell Biol. 2003; 5:59–63.
Article12. Kim S, Cho W, Kim I, Lee SH, Oh GT, Park YM. Oxidized LDL induces vimentin secretion by macrophages and contributes to atherosclerotic inflammation. J Mol Med (Berl). 2020; 98:973–83.
Article13. Gong DH, Dai Y, Chen S, Wang XQ, Yan XX, Shen Y, et al. Secretory vimentin is associated with coronary artery disease in patients and induces atherogenesis in ApoE-/- mice. Int J Cardiol. 2019; 283:9–16.
Article14. Jay AG, Chen AN, Paz MA, Hung JP, Hamilton JA. CD36 binds oxidized low density lipoprotein (LDL) in a mechanism dependent upon fatty acid binding. J Biol Chem. 2015; 290:4590–603.
Article15. Eriksson JE, He T, Trejo-Skalli AV, Harmala-Brasken AS, Hellman J, Chou YH, et al. Specific in vivo phosphorylation sites determine the assembly dynamics of vimentin intermediate filaments. J Cell Sci. 2004; 117(Pt 6):919–32.
Article16. Kim SY, Jeong SJ, Park JH, Cho W, Ahn YH, Choi YH, et al. Plasma membrane localization of CD36 requires vimentin phosphorylation; a mechanism by which macrophage vimentin promotes atherosclerosis. Front Cardiovasc Med. 2022; 9:792717.
Article17. Rea S, James DE. Moving GLUT4: the biogenesis and trafficking of GLUT4 storage vesicles. Diabetes. 1997; 46:1667–77.
Article18. Wieman HL, Wofford JA, Rathmell JC. Cytokine stimulation promotes glucose uptake via phosphatidylinositol-3 kinase/Akt regulation of Glut1 activity and trafficking. Mol Biol Cell. 2007; 18:1437–46.
Article19. Shigyo M, Tohda C. Extracellular vimentin is a novel axonal growth facilitator for functional recovery in spinal cord-injured mice. Sci Rep. 2016; 6:28293.
Article20. Li W, Miller WT. Role of the activation loop tyrosines in regulation of the insulin-like growth factor I receptor-tyrosine kinase. J Biol Chem. 2006; 281:23785–91.
Article21. Montessuit C, Thorburn A. Transcriptional activation of the glucose transporter GLUT1 in ventricular cardiac myocytes by hypertrophic agonists. J Biol Chem. 1999; 274:9006–12.
Article22. Basse AL, Isidor MS, Winther S, Skjoldborg NB, Murholm M, Andersen ES, et al. Regulation of glycolysis in brown adipocytes by HIF-1α. Sci Rep. 2017; 7:4052.
Article23. Reue K, Dwyer JR. Lipin proteins and metabolic homeostasis. J Lipid Res. 2009; 50 Suppl:S109–14.
Article24. Zechner R, Zimmermann R, Eichmann TO, Kohlwein SD, Haemmerle G, Lass A, et al. FAT SIGNALS: lipases and lipolysis in lipid metabolism and signaling. Cell Metab. 2012; 15:279–91.25. Koumenis C, Naczki C, Koritzinsky M, Rastani S, Diehl A, Sonenberg N, et al. Regulation of protein synthesis by hypoxia via activation of the endoplasmic reticulum kinase PERK and phosphorylation of the translation initiation factor eIF2alpha. Mol Cell Biol. 2002; 22:7405–16.26. Ozcan U, Yilmaz E, Ozcan L, Furuhashi M, Vaillancourt E, Smith RO, et al. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science. 2006; 313:1137–40.
Article27. Shu S, Wang H, Zhu J, Liu Z, Yang D, Wu W, et al. Reciprocal regulation between ER stress and autophagy in renal tubular fibrosis and apoptosis. Cell Death Dis. 2021; 12:1016.
Article28. Perez-Sala D, Oeste CL, Martinez AE, Carrasco MJ, Garzon B, Canada FJ. Vimentin filament organization and stress sensing depend on its single cysteine residue and zinc binding. Nat Commun. 2015; 6:7287.
Article29. Rhoads JP, Major AS. How oxidized low-density lipoprotein activates inflammatory responses. Crit Rev Immunol. 2018; 38:333–42.
Article30. Santiago-Fernandez C, Martin-Reyes F, Tome M, Ocana-Wilhelmi L, Rivas-Becerra J, Tatzber F, et al. Oxidized LDL modify the human adipocyte phenotype to an insulin resistant, proinflamatory and proapoptotic profile. Biomolecules. 2020; 10:534.
Article31. Ebeling P, Koistinen HA, Koivisto VA. Insulin-independent glucose transport regulates insulin sensitivity. FEBS Lett. 1998; 436:301–3.
Article32. Hwang DY, Ismail-Beigi F. Stimulation of GLUT-1 glucose transporter expression in response to hyperosmolarity. Am J Physiol Cell Physiol. 2001; 281:C1365–72.
Article33. Beg M, Abdullah N, Thowfeik FS, Altorki NK, McGraw TE. Distinct Akt phosphorylation states are required for insulin regulated Glut4 and Glut1-mediated glucose uptake. Elife. 2017; 6:e26896.
Article34. Feng J, Han J, Pearce SF, Silverstein RL, Gotto AM Jr, Hajjar DP, et al. Induction of CD36 expression by oxidized LDL and IL-4 by a common signaling pathway dependent on protein kinase C and PPAR-gamma. J Lipid Res. 2000; 41:688–96.35. Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science. 1993; 259:87–91.
Article36. McGowan KM, Police S, Winslow JB, Pekala PH. Tumor necrosis factor-alpha regulation of glucose transporter (GLUT1) mRNA turnover: contribution of the 3’-untranslated region of the GLUT1 message. J Biol Chem. 1997; 272:1331–7.37. Cornelius P, Marlowe M, Lee MD, Pekala PH. The growth factor-like effects of tumor necrosis factor-alpha. Stimulation of glucose transport activity and induction of glucose transporter and immediate early gene expression in 3T3-L1 preadipocytes. J Biol Chem. 1990; 265:20506–16.
Article38. Bird TA, Davies A, Baldwin SA, Saklatvala J. Interleukin 1 stimulates hexose transport in fibroblasts by increasing the expression of glucose transporters. J Biol Chem. 1990; 265:13578–83.
Article39. Pendergrass M, Koval J, Vogt C, Yki-Jarvinen H, Iozzo P, Pipek R, et al. Insulin-induced hexokinase II expression is reduced in obesity and NIDDM. Diabetes. 1998; 47:387–94.
Article40. Laufs U, Parhofer KG, Ginsberg HN, Hegele RA. Clinical review on triglycerides. Eur Heart J. 2020; 41:99–109. c.
Article41. Karpe F, Dickmann JR, Frayn KN. Fatty acids, obesity, and insulin resistance: time for a reevaluation. Diabetes. 2011; 60:2441–9.
Article42. Baldini F, Fabbri R, Eberhagen C, Voci A, Portincasa P, Zischka H, et al. Adipocyte hypertrophy parallels alterations of mitochondrial status in a cell model for adipose tissue dysfunction in obesity. Life Sci. 2021; 265:118812.
Article43. He Q, Gao Z, Yin J, Zhang J, Yun Z, Ye J. Regulation of HIF-1 {alpha} activity in adipose tissue by obesity-associated factors: adipogenesis, insulin, and hypoxia. Am J Physiol Endocrinol Metab. 2011; 300:E877–85.44. Kim JW, Tchernyshyov I, Semenza GL, Dang CV. HIF-1-mediated expression of pyruvate dehydrogenase kinase: a metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006; 3:177–85.
Article45. Kierans SJ, Taylor CT. Regulation of glycolysis by the hypoxiainducible factor (HIF): implications for cellular physiology. J Physiol. 2021; 599:23–37.
Article46. Chen C, Pore N, Behrooz A, Ismail-Beigi F, Maity A. Regulation of glut1 mRNA by hypoxia-inducible factor-1: interaction between H-ras and hypoxia. J Biol Chem. 2001; 276:9519–25.47. Zheng Z, Zhang C, Zhang K. Role of unfolded protein response in lipogenesis. World J Hepatol. 2010; 2:203–7.
Article48. Takahashi N, Yoshizaki T, Hiranaka N, Suzuki T, Yui T, Akanuma M, et al. Endoplasmic reticulum stress suppresses lipin-1 expression in 3T3-L1 adipocytes. Biochem Biophys Res Commun. 2013; 431:25–30.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Adipocyte Signals in Energy Balance and Digestive Diseases
- The Role of Vitamin D in Adipose Tissue Biology: Adipocyte Differentiation, Energy Metabolism, and Inflammation
- Brown Fat and Browning for the Treatment of Obesity and Related Metabolic Disorders
- Transcriptional repression of type I procollagen genes during adipocyte differentiation
- The Mechanism of White and Brown Adipocyte Differentiation
