Insulin Preferentially Regulates the Activity of Parasympathetic Preganglionic Neurons over Sympathetic Preganglionic Neurons
- Affiliations
-
- 1Department of Biological Sciences, Korea Advanced Institute of Science and Technology, Daejeon, Korea
- KMID: 2546985
- DOI: http://doi.org/10.3803/EnM.2023.1725
Abstract
- Background
Insulin is a peptide hormone that regulates post-prandial physiology, and it is well known that insulin controls homeostasis at least in part via the central nervous system. In particular, insulin alters the activity of neurons within the autonomic nervous system. However, currently available data are mostly from unidentified brainstem neurons of the dorsal motor nucleus of the vagus nerve (DMV).
Methods
In this study, we used several genetically engineered mouse models to label distinct populations of neurons within the brainstem and the spinal cord for whole-cell patch clamp recordings and to assess several in vivo metabolic functions.
Results
We first confirmed that insulin directly inhibited cholinergic (parasympathetic preganglionic) neurons in the DMV. We also found inhibitory effects of insulin on both the excitatory and inhibitory postsynaptic currents recorded in DMV cholinergic neurons. In addition, GABAergic neurons of the DMV and nucleus tractus solitarius were inhibited by insulin. However, insulin had no effects on the cholinergic sympathetic preganglionic neurons of the spinal cord. Finally, we obtained results suggesting that the insulininduced inhibition of parasympathetic preganglionic neurons may not play a critical role in the regulation of glucose homeostasis and gastrointestinal motility.
Conclusion
Our results demonstrate that insulin inhibits parasympathetic neuronal circuitry in the brainstem, while not affecting sympathetic neuronal activity in the spinal cord.
Keyword
Figure
-
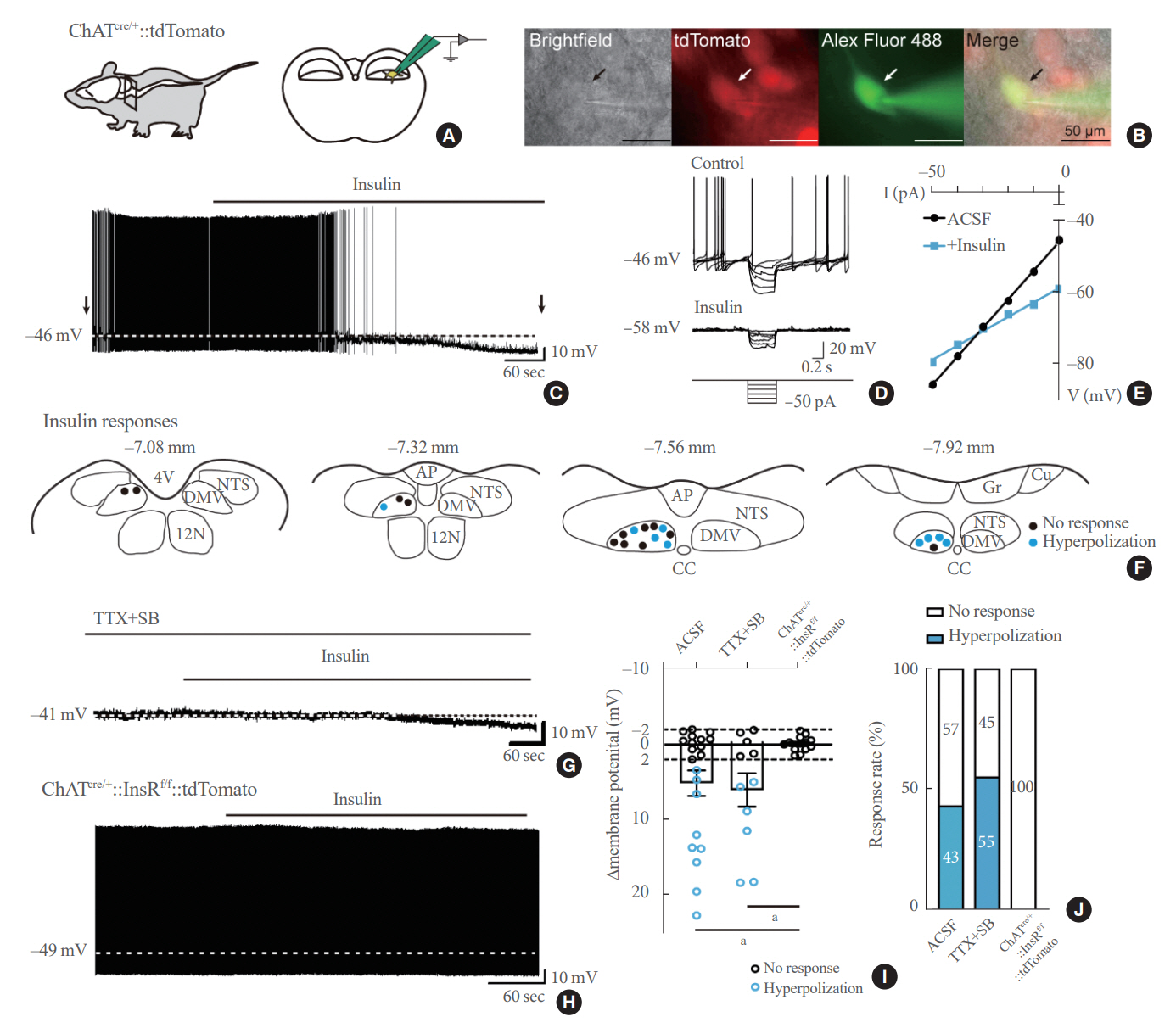
Fig. 1. Insulin directly inhibits parasympathetic preganglionic neurons. (A) Image demonstrating the location of the brainstem section (left) and a parasympathetic preganglionic neuron of the dorsal motor nucleus of the vagus nerve (DMV; right) targeted for whole-cell patch clamp recordings, obtained from ChATcre/+::tdTomato mice. (B) Bright-field, tetramethylrhodamine (TRITC), fluorescein isothiocyanate (FITC), and merged images (from left to right) of the targeted neuron (arrows) in the DMV of ChATcre/+::tdTomato mice. Scale bar=50 μm. (C) Membrane potential hyperpolarizes in response to insulin (50 nM) treatment. The dashed line indicates the baseline membrane potential. (D) Voltage responses to hyperpolarizing current steps from –50 to 0 pA, applied as indicated by arrows in (C). (E) The voltage-current relationship demonstrates insulin-induced decreases in input resistance. (F) Drawings of four rostrocaudal levels of the mouse brainstem summarize the location of the acute effects of insulin on parasympathetic preganglionic neurons. Blue dots indicate hyperpolarized cells, while black dots indicate cells with no effects. (G) The membrane potential hyperpolarized in response to insulin treatment in the presence of tetrodotoxin (TTX, 500 nM) and synaptic blockers (SBs, 1 mM kynurenic acid+50 μM picrotoxin). The dashed line indicates baseline membrane potential. (H) The membrane potential did not change after insulin treatment parasympathetic preganglionic neurons from ChATcre/+::InsRf/f::tdTomato mice. The dashed line indicates baseline membrane potential. (I) Plots summarizing insulin-induced changes in membrane potential. (J) Histogram summarizing the percentage of neurons showing insulin responses. ACSF, artificial cerebrospinal fluid; 4V, fourth ventricle; 12N, nucleus of the hypoglossal nerve; NTS, nucleus tractus solitaries; AP, area postrema; CC, central canal; Cu, nucleus cuneatus; Gr, nucleus gracilis; ChAT, choline acetyltransferase; InsRf/f, insulin receptor flox. aP<0.01, Fisher’s exact test for the occurrence of hyperpolarization and no effects.
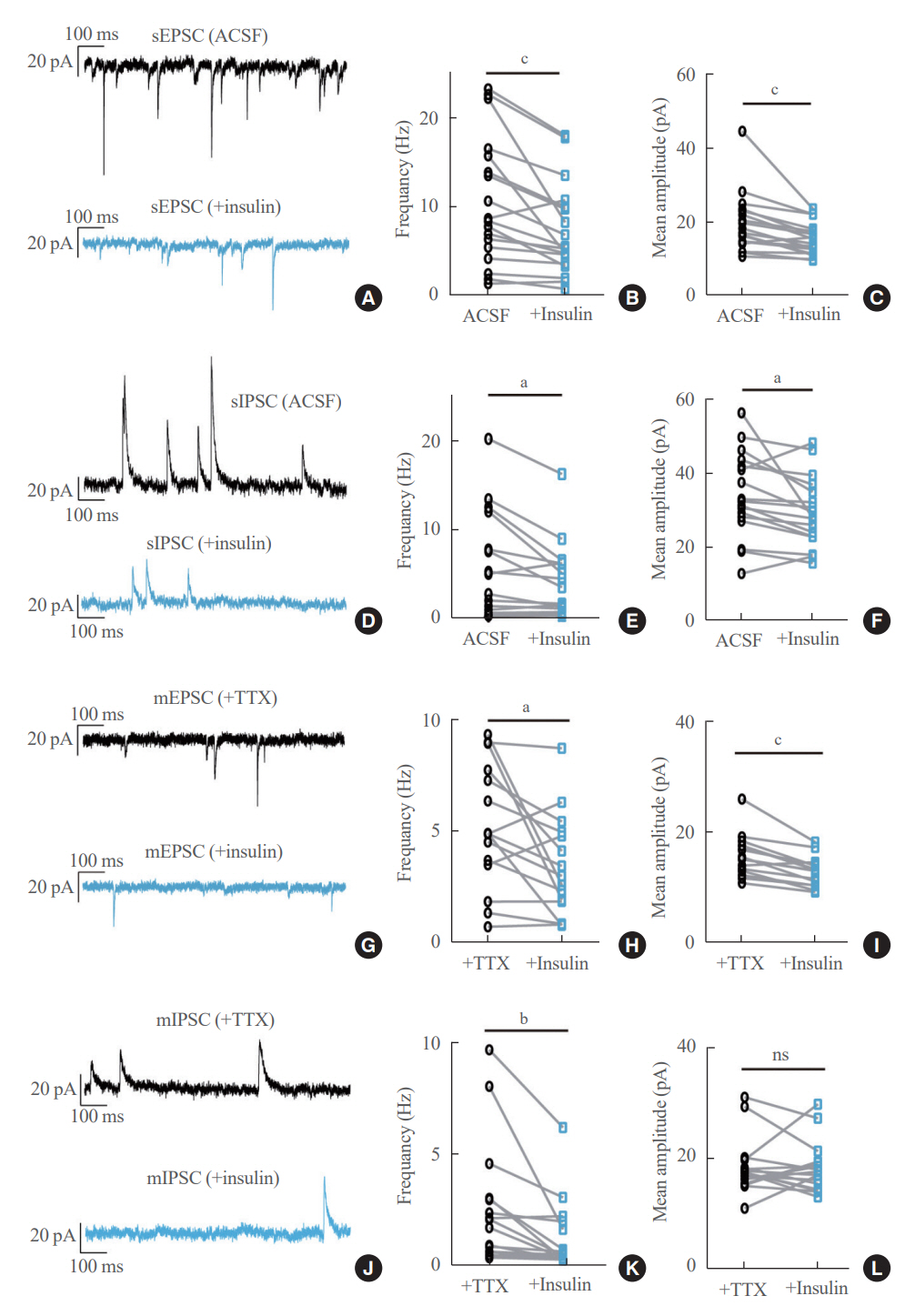
Fig. 2. Insulin reduces excitatory and inhibitory postsynaptic currents onto parasympathetic preganglionic neurons. (A) Traces demonstrating spontaneous excitatory postsynaptic currents (sEPSCs) before (upper) and during (lower) insulin (50 nM) treatment. Holding potential=–60 mV. (B, C) Insulin reduced the frequency (B) and mean amplitude (C) of sEPSCs. (D) Traces demonstrating spontaneous inhibitory postsynaptic currents (sIPSCs) before (upper) and during (lower) insulin treatment. Holding potential=–10 mV. (E, F) Insulin reduced the frequency (E) and mean amplitude (F) of sIPSCs. (G) Traces demonstrating miniature EPSCs (mEPSCs) before (upper) and during (lower) insulin treatment. Holding potential=–60 mV. (H, I) Insulin reduced the frequency (H) and mean amplitude (I) of mEPSCs. (J) Traces demonstrating miniature IPSCs (mIPSCs) before (upper) and during (lower) insulin treatment. Holding potential=–10 mV. (K, L) Insulin reduced the frequency (K), but not the mean amplitude (L) of mIPSCs. ACSF, artificial cerebrospinal fluid; TTX, tetrodotoxin; ns, not significant (P>0.05). aP<0.05; bP<0.01; cP<0.001 by the Wilcoxon matched-pairs signed rank test.

Fig. 3. Insulin hyperpolarizes GABAergic neurons of the nucleus tractus solitaries (NTS) and dorsal motor nucleus of the vagus nerve (DMV). (A) Bright-field, tetramethylrhodamine (TRITC), fluorescein isothiocyanate (FITC), and merged images (from left to right) of the targeted neuron (arrows) in the NTS of Phox2bcre/+::tdTomato mice. Scale bar=50 μm. (B) The membrane potential hyperpolarized in response to insulin (50 nM) treatment. The dashed line indicates the baseline membrane potential. (C) Plots summarizing insulin-induced changes in membrane potential. (D) Bright-field, TRITC, FITC, and merged images (from left to right) of the targeted neuron (arrows) in the NTS of Vgatcre/+::tdTomato mice. Scale bar=50 μm. (E) The membrane potential hyperpolarized in response to insulin treatment. The dashed line indicates the baseline membrane potential. (F) Plots summarize insulin-induced changes in the membrane potential. (G) Bright-field, TRITC, FITC, and merged images (from left to right) of the targeted neuron (arrows) in the DMV of Vgatcre/+::tdTomato mice. Scale bar=50 μm. (H) The membrane potential hyperpolarized in response to insulin treatment. The dashed line indicates the baseline membrane potential. (I) Plots summarizing insulin-induced changes in the membrane potential. (J) Illustration summarizing modulation of parasympathetic preganglionic neurons and their upstream neurons by insulin and insulin receptors. Phox2b, paired-like homeobox 2b; Vgat, vesicular GABA transporter; ACSF, artificial cerebrospinal fluid; AP, area postrema; ChAT, choline acetyltransferase; InsR, insulin receptor; GABA, γ-aminobutyric acid.
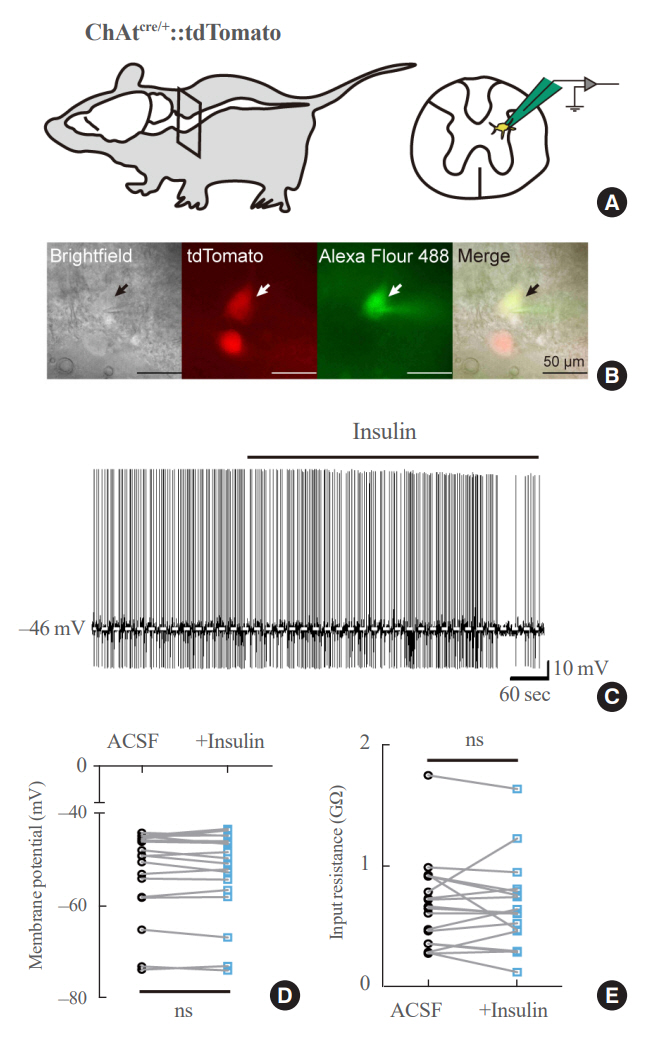
Fig. 4. Insulin does not affect the activity of sympathetic preganglionic neurons. (A) Image demonstrating the location of the spinal cord section (left) and the sympathetic preganglionic neuron of intermediolateral column (IML, right) targeted for whole-cell patch clamp recordings, obtained from ChATcre/+::tdTomato mice. (B) Bright-field, tetramethylrhodamine (TRITC), fluorescein isothiocyanate (FITC), and merged images (from left to right) of targeted neuron (arrows) in the IML of ChATcre/+::tdTomato mice. Scale bar=50 μm. (C) The membrane potential did not change in response to insulin (50 nM) treatment. The dashed line indicates the baseline membrane potential. (D, E) Plots summarizing insulin-induced changes in the membrane potential (D) and input resistance (E). Not significant (ns) (P>0.05) by Wilcoxon matched-pairs signed rank test. ChAT, choline acetyltransferase; ACSF, artificial cerebrospinal fluid.

Reference
-
1. Chen C, Cohrs CM, Stertmann J, Bozsak R, Speier S. Human beta cell mass and function in diabetes: recent advances in knowledge and technologies to understand disease pathogenesis. Mol Metab. 2017; 6:943–57.
Article2. Sonksen P, Sonksen J. Insulin: understanding its action in health and disease. Br J Anaesth. 2000; 85:69–79.
Article3. Brüning JC, Gautam D, Burks DJ, Gillette J, Schubert M, Orban PC, et al. Role of brain insulin receptor in control of body weight and reproduction. Science. 2000; 289:2122–5.
Article4. Klockener T, Hess S, Belgardt BF, Paeger L, Verhagen LA, Husch A, et al. High-fat feeding promotes obesity via insulin receptor/PI3K-dependent inhibition of SF-1 VMH neurons. Nat Neurosci. 2011; 14:911–8.
Article5. Konner AC, Janoschek R, Plum L, Jordan SD, Rother E, Ma X, et al. Insulin action in AgRP-expressing neurons is required for suppression of hepatic glucose production. Cell Metab. 2007; 5:438–49.
Article6. Hill JW, Elias CF, Fukuda M, Williams KW, Berglund ED, Holland WL, et al. Direct insulin and leptin action on proopiomelanocortin neurons is required for normal glucose homeostasis and fertility. Cell Metab. 2010; 11:286–97.
Article7. Filippi BM, Yang CS, Tang C, Lam TK. Insulin activates Erk1/2 signaling in the dorsal vagal complex to inhibit glucose production. Cell Metab. 2012; 16:500–10.
Article8. Filippi BM, Bassiri A, Abraham MA, Duca FA, Yue JT, Lam TK. Insulin signals through the dorsal vagal complex to regulate energy balance. Diabetes. 2014; 63:892–9.
Article9. Blake CB, Smith BN. Insulin reduces excitation in gastricrelated neurons of the dorsal motor nucleus of the vagus. Am J Physiol Regul Integr Comp Physiol. 2012; 303:R807–14.
Article10. Grabauskas G, Moises HC. Gastrointestinal-projecting neurones in the dorsal motor nucleus of the vagus exhibit direct and viscerotopically organized sensitivity to orexin. J Physiol. 2003; 549(Pt 1):37–56.
Article11. Cork SC, Richards JE, Holt MK, Gribble FM, Reimann F, Trapp S. Distribution and characterisation of glucagon-like peptide-1 receptor expressing cells in the mouse brain. Mol Metab. 2015; 4:718–31.
Article12. Zheng Z, Lewis MW, Travagli RA. In vitro analysis of the effects of cholecystokinin on rat brain stem motoneurons. Am J Physiol Gastrointest Liver Physiol. 2005; 288:G1066–73.
Article13. Sohn JW, Harris LE, Berglund ED, Liu T, Vong L, Lowell BB, et al. Melanocortin 4 receptors reciprocally regulate sympathetic and parasympathetic preganglionic neurons. Cell. 2013; 152:612–9.
Article14. Ju SH, Yun H, Oh Y, Choi Y, Sohn JW. Melanocortin-4 receptors activate sympathetic preganglionic neurons and elevate blood pressure via TRPV1. Cell Rep. 2022; 41:111579.
Article15. Chambers AP, Smith EP, Begg DP, Grayson BE, Sisley S, Greer T, et al. Regulation of gastric emptying rate and its role in nutrient-induced GLP-1 secretion in rats after vertical sleeve gastrectomy. Am J Physiol Endocrinol Metab. 2014; 306:E424–32.
Article16. Rossi J, Balthasar N, Olson D, Scott M, Berglund E, Lee CE, et al. Melanocortin-4 receptors expressed by cholinergic neurons regulate energy balance and glucose homeostasis. Cell Metab. 2011; 13:195–204.
Article17. Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci. 2010; 13:133–40.
Article18. Bruning JC, Michael MD, Winnay JN, Hayashi T, Horsch D, Accili D, et al. A muscle-specific insulin receptor knockout exhibits features of the metabolic syndrome of NIDDM without altering glucose tolerance. Mol Cell. 1998; 2:559–69.
Article19. Kang BJ, Chang DA, Mackay DD, West GH, Moreira TS, Takakura AC, et al. Central nervous system distribution of the transcription factor Phox2b in the adult rat. J Comp Neurol. 2007; 503:627–41.
Article20. Davis SF, Derbenev AV, Williams KW, Glatzer NR, Smith BN. Excitatory and inhibitory local circuit input to the rat dorsal motor nucleus of the vagus originating from the nucleus tractus solitarius. Brain Res. 2004; 1017:208–17.
Article21. Scott MM, Williams KW, Rossi J, Lee CE, Elmquist JK. Leptin receptor expression in hindbrain Glp-1 neurons regulates food intake and energy balance in mice. J Clin Invest. 2011; 121:2413–21.
Article22. Vong L, Ye C, Yang Z, Choi B, Chua S Jr, Lowell BB. Leptin action on GABAergic neurons prevents obesity and reduces inhibitory tone to POMC neurons. Neuron. 2011; 71:142–54.
Article23. Hyun U, Sohn JW. Autonomic control of energy balance and glucose homeostasis. Exp Mol Med. 2022; 54:370–6.
Article24. Kwon E, Joung HY, Liu SM, Chua SC Jr, Schwartz GJ, Jo YH. Optogenetic stimulation of the liver-projecting melanocortinergic pathway promotes hepatic glucose production. Nat Commun. 2020; 11:6295.
Article25. Yang F, Liu Y, Chen S, Dai Z, Yang D, Gao D, et al. A GABAergic neural circuit in the ventromedial hypothalamus mediates chronic stress-induced bone loss. J Clin Invest. 2020; 130:6539–54.
Article26. Sohn JW, Oh Y, Kim KW, Lee S, Williams KW, Elmquist JK. Leptin and insulin engage specific PI3K subunits in hypothalamic SF1 neurons. Mol Metab. 2016; 5:669–79.
Article27. Williams KW, Margatho LO, Lee CE, Choi M, Lee S, Scott MM, et al. Segregation of acute leptin and insulin effects in distinct populations of arcuate proopiomelanocortin neurons. J Neurosci. 2010; 30:2472–9.
Article28. Card JP, Enquist LW. Transneuronal circuit analysis with pseudorabies viruses. Curr Protoc Neurosci. 2014; 68:1.5.1–1.5.39.
Article29. Zhou SY, Lu YX, Yao H, Owyang C. Spatial organization of neurons in the dorsal motor nucleus of the vagus synapsing with intragastric cholinergic and nitric oxide/VIP neurons in the rat. Am J Physiol Gastrointest Liver Physiol. 2008; 294:G1201–9.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Neurophysiology of Laryngopharyngeal Reflux and Brainstem Reflex
- The Frequency-Dependence of Pre- and Postganglionic Nerve Stimulation of Pig and Rat Bladder
- Sympathetic Excitation of Afferent Neurons within Dorsal Root Ganglia in a Rat Model of Sympathetically Medicated Pain
- Diagnostic Usefulness of Electrodiagnostic Study and Magnetic Resonance Imaging to Preganglionic Brachial Plexopathy
- Identification of ATP-sensitive K+ Conductances in Male Rat Major Pelvic Ganglion Neurons
