Hyperglycemia-Suppressed SMARCA5 Disrupts Transcriptional Homeostasis to Facilitate Endothelial Dysfunction in Diabetes
- Affiliations
-
- 1Departments of Nursing, Xi’an No. 5 Hospital, Xi’an, China
- 2Departments of Geriatric, Xi’an No. 5 Hospital, Xi’an, China
- KMID: 2542514
- DOI: http://doi.org/10.4093/dmj.2022.0179
Abstract
- Background
Dysfunction of vascular endothelial cells (ECs) plays a central role in the pathogenesis of cardiovascular complications in diabetes. SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) is a key regulator of chromatin structure and DNA repair, but its role in ECs remains surprisingly unexplored. The current study was designed to elucidate the regulated expression and function of SMARCA5 in diabetic ECs.
Methods
SMARCA5 expression was evaluated in ECs from diabetic mouse and human circulating CD34+ cells using quantitative reverse transcription polymerase chain reaction and Western blot. Effects of SMARCA5 manipulation on ECs function were evaluated using cell migration, in vitro tube formation and in vivo wound healing assays. Interaction among oxidative stress, SMARCA5 and transcriptional reprogramming was elucidated using luciferase reporter assay, electrophoretic mobility shift assay and chromatin immunoprecipitation.
Results
Endothelial SMARCA5 expression was significantly decreased in diabetic rodents and humans. Hyperglycemia-suppressed SMARCA5 impaired EC migration and tube formation in vitro, and blunted vasculogenesis in vivo. Contrarily, overexpression of SMARCA5 in situ by a SMARCA5 adenovirus-incorporated hydrogel effectively promoted the rate of wound healing in a dorsal skin punch injury model of diabetic mice. Mechanistically, hyperglycemia-elicited oxidative stress suppressed SMARCA5 transactivation in a signal transducer and activator of transcription 3 (STAT3)-dependent manner. Moreover, SMARCA5 maintained transcriptional homeostasis of several pro-angiogenic factors through both direct and indirect chromatin-remodeling mechanisms. In contrast, depletion of SMARCA5 disrupted transcriptional homeostasis to render ECs unresponsive to established angiogenic factors, which ultimately resulted in endothelial dysfunction in diabetes.
Conclusion
Suppression of endothelial SMARCA5 contributes to, at least in part, multiple aspects of endothelial dysfunction, which may thereby exacerbate cardiovascular complications in diabetes.
Keyword
Figure
-
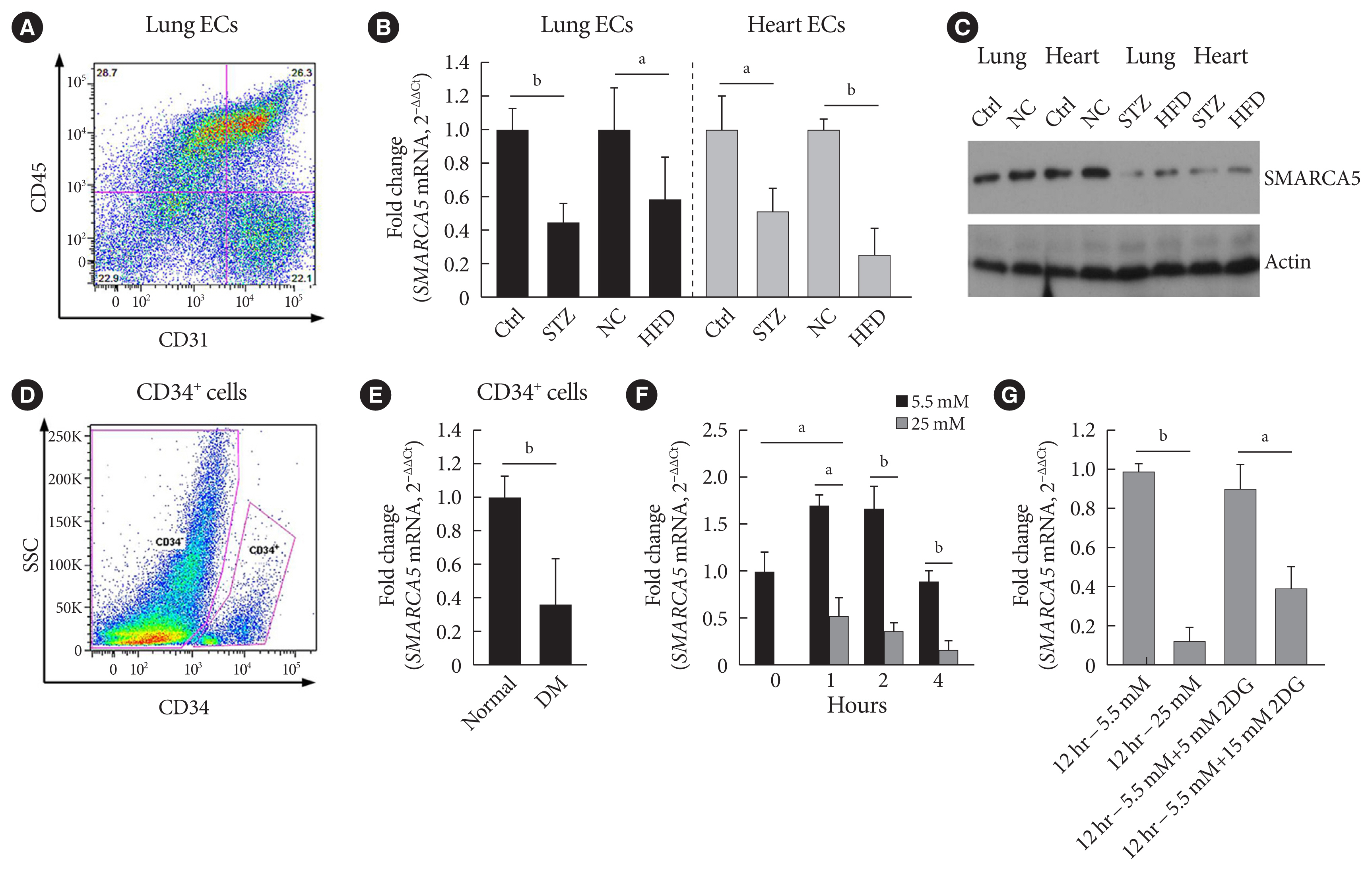
Fig. 1 Diabetes attenuates SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) expression in endothelial cells (ECs). (A) Representative fluorescence activated cell sorting (FACS) plots for sorting ECs from lung based on sorting live/CD45−/CD31+ cells. (B) Relative expression levels of Smarca5 mRNA in different ECs were evaluated and quantified using quantitative polymerase chain reaction (qPCR) (n=4). Amplification of 18S was used to obtain the ΔΔCt values for the calculation of fold changes. (C) Western blot analysis of SMARCA5 protein expression in different ECs. β-Actin served as loading control (Ctrl). (D) Human CD34+ cells were isolated from the peripheral blood samples of type 2 diabetes mellitus (T2DM) patients and normal volunteers using FACS method. (E) Relative SMARCA5 mRNA abundance in vasculogenic circulating CD34+ cells isolated from patients with T2DM, versus matched normal subjects, was determined by qPCR (n=4). (F) Relative Smarca5 mRNA abundance in lung ECs after exposing cells to hyperglycemia was assessed using qPCR (n=4). (G) Relative Smarca5 mRNA abundance in lung ECs in absence or presence of 2-deoxyglucose (2DG) for 12 hours was assessed using qPCR (n=4). Results presented as mean±standard deviation of three independent experiments. STZ, streptozotocin; NC, negative control; HFD, high-fat diet; SSC, side scatter light; DM, diabetes mellitus. aP<0.05, bP<0.01.
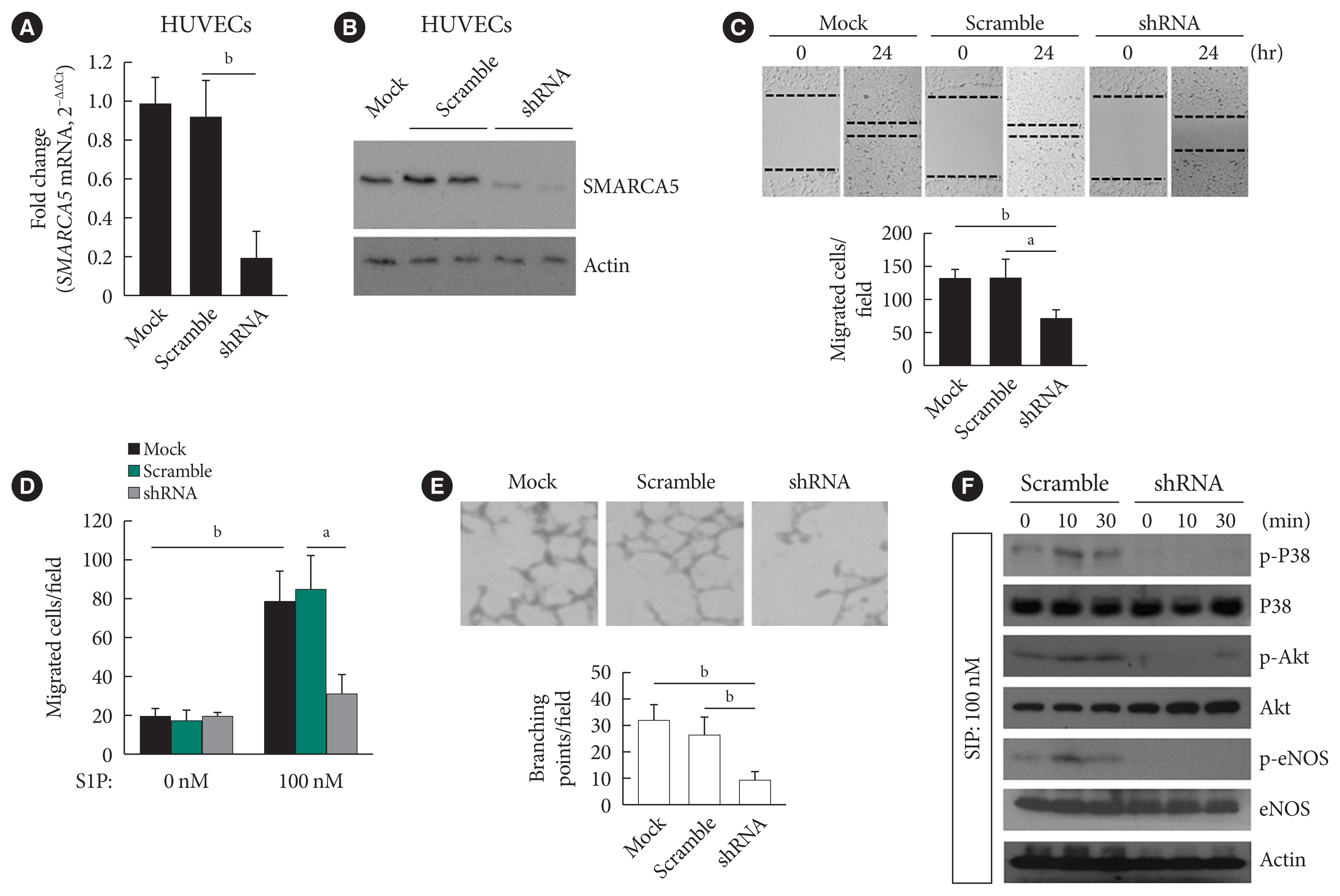
Fig. 2 Ablation of SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) impairs endothelial function. (A) Stable knockdown of SMARCA5 in human umbilical vein endothelial cells (HUVECs) was verified using quantitative polymerase chain reaction. (B) Stable knockdown of SMARCA5 in HUVECs was verified at protein level using Western blot. (C) Stable inhibition of SMARCA5 suppressed migration, as compared to mock or scramble shRNA-treated HUVECs, in scratch assays (n=5). (D) Stable inhibition of SMARCA5 suppressed migration, as compared to mock or scramble shRNA-treated HUVECs, in Transwell assays stimulated by 100 nM of sphingosine 1-phosphate (S1P) for 12 hours (n=5). (E) Stable knockdown of SMARCA5 inhibited the tube-forming activity of HUVECs (n=5). (F) HUVECsSMARCA5−/− or Scramble shRNA-treated HUVECs were incubated with 100 nM of S1P for different durations as indicated, followed by Western blot analysis of activation of P38, Akt, and endothelial nitric oxide synthase (eNOS) using corresponding phospho-specific antibodies. aP<0.05, bP<0.01.
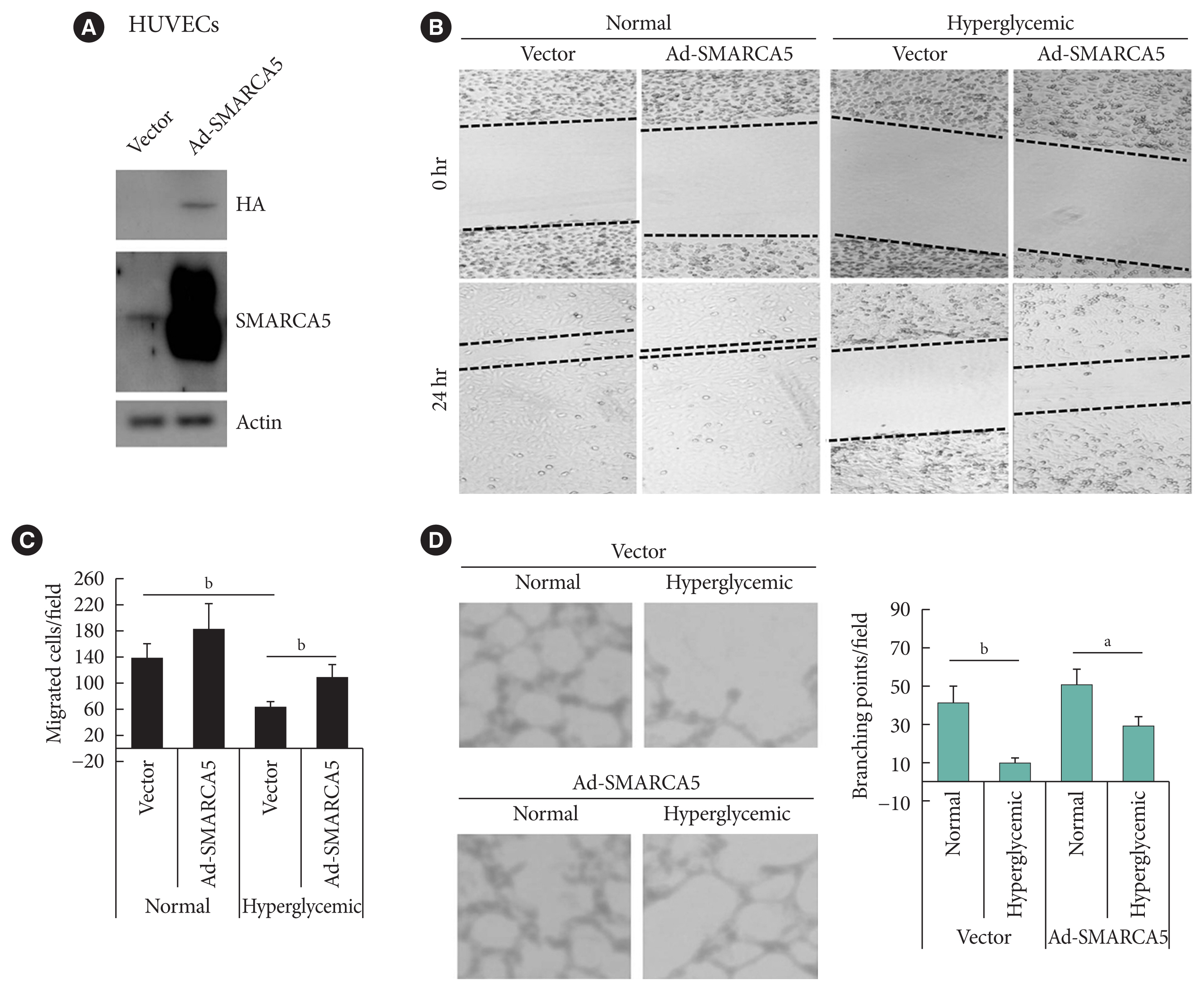
Fig. 3 Ectopic overexpression of SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) ameliorates endothelial dysfunction in vitro. (A) Stable overexpression of SMARCA5 in human umbilical vein endothelial cells (HUVECs) was verified using Western blot. (B, C) Stable SMARCA5 overexpression improved migration under hyperglycemic conditions (incubation with 25 mM glucose for 24), as compared to vector-treated HUVECs, in scratch assays (n=5). (D) Stable SMARCA5 overexpression promoted the tube-forming activity of HUVECs under hyperglycemic conditions (incubation with 25 mM glucose for 24) (n=5). HA, hemagglutinin. aP<0.05, bP<0.01.
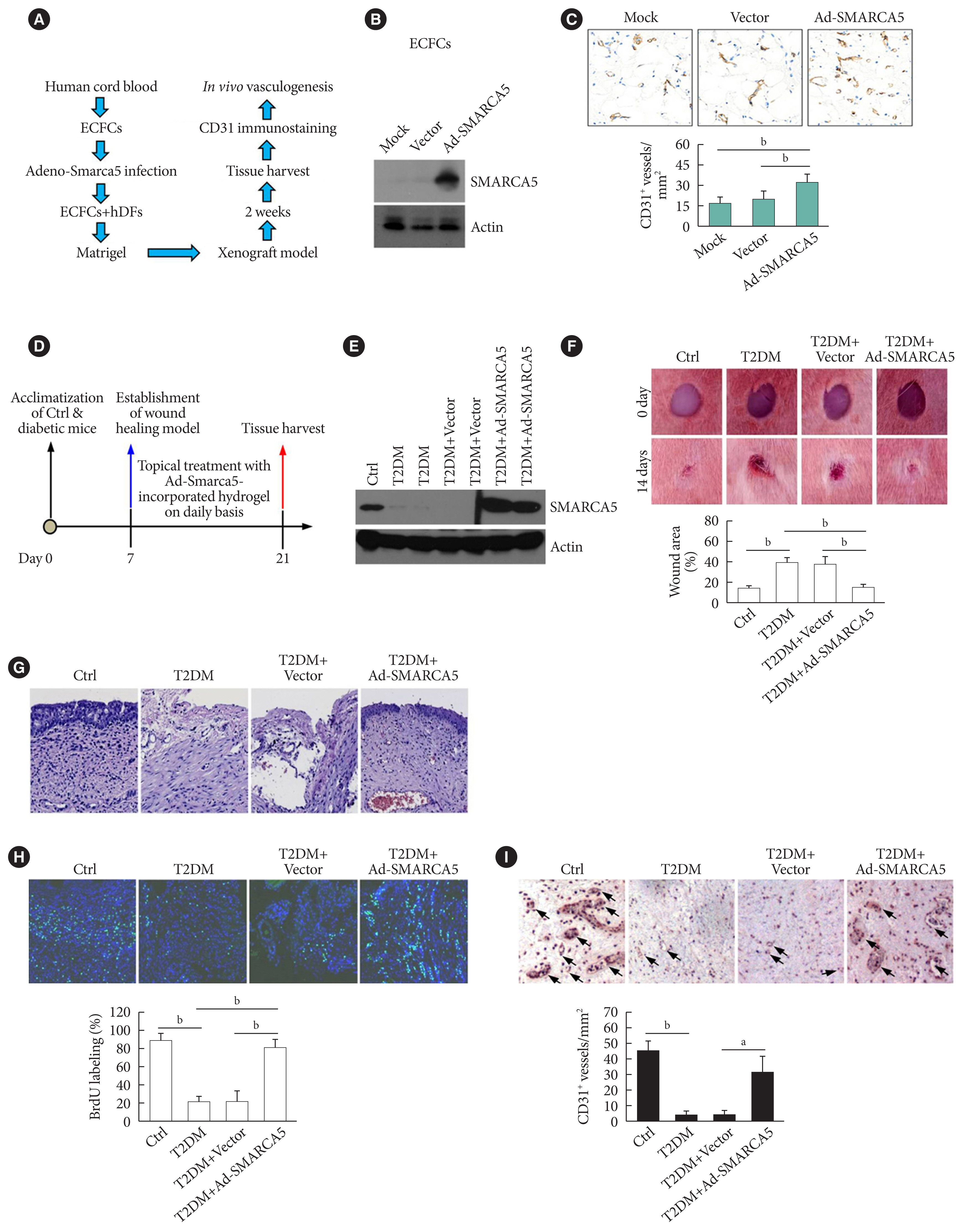
Fig. 4 Induced expression of endothelial SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) in vivo ameliorates diabetic vascular dysfunction. (A) Diagram of procedures employed in vasculogenesis assay in vivo. (B) Effects of adenovirus overexpression of exogenous SMARCA5 in human endothelial colony forming cells (ECFCs) were confirmed by Western blot. (C) In vivo vasculogenesis assay using human dermal fibroblast (hDFs) and human ECFCs expressing empty vector versus Ad-SMARCA5. Final vasculogenesis in vivo was evaluated by measuring numbers of CD31+ vessels per analyzed areas (mm2) (n=5). (D) Diagram of procedures employed in overexpression of SMARCA5 in situ and in vivo wound healing assay. (E) At 14 days after topical treatment with Ad-Smarca5 incorporated hydrogel, subcutaneous tissue homogenates from different experimental mice were subjected to Western blot to analyze SMARCA5 expression. (F) Representative images and quantification of skin wound healing in Ad-Smarca5 incorporated hydrogel-treated diabetic animals. Wound area measurements were conducted using ImageJ software (n=5). (G) 4% Paraformaldehyde-fixed paraffin-embedded skin sections (5 μm) were subjected to haematoxylin and eosin (H&E) staining to explore pathological changes (bar=50 μm). (H) Six days after establishment of wound healing model, mice were injected intraperitoneally with 50 mg/kg 5-bromodeoxyuridine (BrdU) on a daily basis. At 14 days after establishment of wound healing model, mice were sacrificed, and skin tissues were harvested and subjected to BrdU immunofluorescence staining. Quantification of BrdU positive (BrdU+) cells in subcutaneous tissues was carried out with the aid of ImageJ software (bP<0.01, n=5) (bar=50 μm). (I) Subcutaneous vasculogenesis was evaluated by measuring numbers of CD31+ vessels per analyzed areas (mm2) (n=5). Ctrl, control; T2DM, type 2 diabetes mellitus. aP<0.05, bP<0.01.
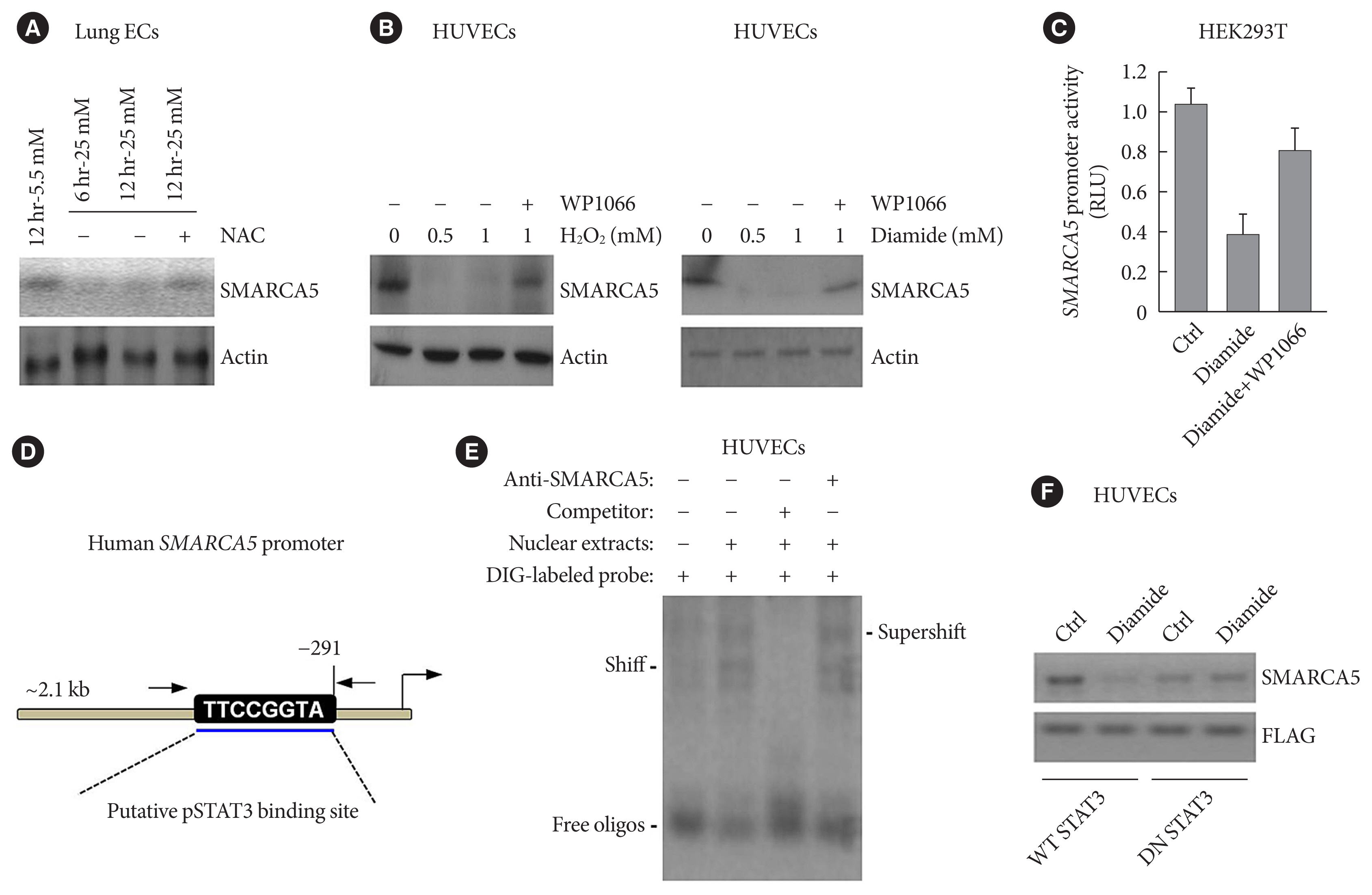
Fig. 5 Oxidative stress regulates SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) expression in a signal transducer and activator of transcription 3 (STAT3)-dependent manner. (A) Primary lung endothelial cells (ECs) were treated with different doses of glucose, in the presence or absence of co-treatment with the ROS scavenger-N-acetyl-L-cysteine (NAC; 10 mM), for 6 or 12 hours, followed by immunoblotting analysis. (B) Human umbilical vein endothelial cells (HUVECs) were treated with different doses of H2O2 or diamide, in the presence or absence of co-treatment with WP1066 (5 μM), for 12 hours, followed by immunoblotting analysis. (C) HEK293T cells were transiently transfected with pGL4-SMARCA5-Luc, as well as the thymidine kinase promoter-renilla luciferase reporter plasmid (pRL-TK) Renilla reporter plasmid. Forty-eight hours later, cells were treated with diamide (1 mM) or diamide plus WP1066 (5 μM) for 12 hours, followed by the measurement of relative luciferase activity. (D) Schematic of a consensus binding site of STAT3 in the SMARCA5 promoter. (E) Electrophoretic mobility shift assay (EMSA) was performed using nuclear extract proteins and a digoxigenin (DIG)-labeled probe containing the consensus pSTAT3 binding sequence. Competitive inhibition consisted of incubation with a 50-fold molar excess of unlabeled/cold probe. Anti-pSTAT3 antibody was also used to confirm specificity of the band. (F) Western blot analysis of SMARCA5 in control (Ctrl) or diamide-treated HUVECs transfected with either wild-type (WT) STAT3 or dominant-negative (DN) STAT3. The FLAG (DYKDDDDK) antibody recognized the transfected WT STAT3 or DN STAT3 in HUVECs. RLU, renilla luciferase activity.

Fig. 6 SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) depletion blocks response to pro-angiogenic cues to inhibit endothelial function. (A) Human umbilical vein endothelial cells (HUVECs)SMARCA5−/− or scramble shRNA-treated HUVECs were incubated with 100 nM of sphingosine 1-phosphate (S1P) for 12 hours, followed by quantitative polymerase chain reaction analysis of expression of pro-angiogenic genes. Results presented as mean±SD of three independent experiments (n=4). The expression of a target mRNA were normalized against the values in Scramble shRNA-treated HUVECs of resting state, which was arbitrarily set at 1 (blue dashed line). (B) Vascular endothelial growth factor (VEGF) concentration in the culture supernatants of HUVECsSMARCA5−/− or Scramble shRNA-treated HUVECs subjected to S1P stimulation (100 nM) for different durations as indicated (n=5). (C) A 0.5 μg pGL4.10-VEGF reporter plasmid and thymidine kinase promoter-renilla luciferase reporter plasmid (pRL-TK) Renilla reporter plasmid were co-transfected into scramble shRNA or SMARCA5 shRNA-transfected HUVECs for. Forty-eight hours later, cells were challenged with 100 nM of S1P for 6 hours, followed by measurement of the relative luciferase activities. (D) Stable overexpression of VEGF in HUVECsSMARCA5−/− was confirmed using Western blot. (E) Stable overexpression of VEGF effectively improved endothelial migration in HUVECsSMARCA5−/−, as compared to vector-transfected HUVECs, in scratch assays (n=5). (F) Stable overexpression of VEGF effectively improved endothelial migration in HUVECsSMARCA5−/−, as compared to vector-transfected HUVECs, in transwell assays stimulated by 100 nM of S1P for 12 hours (n=5). (G) Stable overexpression of VEGF rescued the tube-forming activity of in HUVECsSMARCA5−/− (n=5). Ang-I, angiopoietin-I; bFGF, basic fibroblast growth factor; CTGF, connective tissue growth factor; M-CSF, macrophage colony-stimulating factor; RLU, renilla luciferase activity; pCMV3, the promoter of human cytomegalovirus. aP<0.05, bP<0.01.

Fig. 7 Depletion of SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 (SMARCA5) disrupts the transcriptional homeostasis of pro-angiogenic genes via both direct and indirect chromatin remodeling. (A) Chromatin immunoprecipitation (ChIP) analysis of HUVECs expressing scramble shRNA or SMARCA5 shRNA, in the presence or absence of co-incubation with 100 nM of sphingosine 1-phosphate (S1P) for 6 hours, using antibodies against SMARCA5, Brahma-related gene 1 (BRG1), Brahma homologue (BRM), metastasis-associated protein 1 (MTA1), or enhancer of zeste 2 polycomb repressive complex 2 subunit (EZH2) for the vascular endothelial growth factor (VEGF) gene promoter (n=4). (B) ChIP analysis of HUVECs expressing scramble shRNA or SMARCA5 shRNA, in the presence or absence of co-incubation with 100 nM of S1P for 6 hours, using antibodies against SMARCA5, BRG1, BRM, MTA1, or EZH2 for the basic fibroblast growth factor (bFGF) gene promoter (n=4). (C) A working model depicts the possible mechanisms related to deregulated expression of endothelial SMARCA5 contributing to pathogenesis of the vascular dysfunction by disrupting the transcriptional homeostasis of pro-angiogenic genes in diabetes. UD, undetectable; EC, endothelial cell; Ang-I, angiopoietin-I; eNOS, endothelial nitric oxide synthase; STAT3, signal transducer and activator of transcription 3. aP<0.05, bP<0.01.
Reference
-
1. Knapp M, Tu X, Wu R. Vascular endothelial dysfunction, a major mediator in diabetic cardiomyopathy. Acta Pharmacol Sin. 2019; 40:1–8.2. Yang C, Eleftheriadou M, Kelaini S, Morrison T, Gonzalez MV, Caines R, et al. Targeting QKI-7 in vivo restores endothelial cell function in diabetes. Nat Commun. 2020; 11:3812.3. Aiello LP, Wong JS. Role of vascular endothelial growth factor in diabetic vascular complications. Kidney Int Suppl. 2000; 77:S113–9.4. Kolluru GK, Bir SC, Kevil CG. Endothelial dysfunction and diabetes: effects on angiogenesis, vascular remodeling, and wound healing. Int J Vasc Med. 2012; 2012:918267.
Article5. Menon DU, Kirsanov O, Geyer CB, Magnuson T. Mammalian SWI/SNF chromatin remodeler is essential for reductional meiosis in males. Nat Commun. 2021; 12:6581.6. Thakur S, Cahais V, Turkova T, Zikmund T, Renard C, Stopka T, et al. Chromatin remodeler smarca5 is required for cancer-related processes of primary cell fitness and immortalization. Cells. 2022; 11:808.7. Jevtic Z, Matafora V, Casagrande F, Santoro F, Minucci S, Garre M, et al. SMARCA5 interacts with NUP98-NSD1 oncofusion protein and sustains hematopoietic cells transformation. J Exp Clin Cancer Res. 2022; 41:34.8. Zikmund T, Paszekova H, Kokavec J, Kerbs P, Thakur S, Turkova T, et al. Loss of ISWI ATPase SMARCA5 (SNF2H) in acute myeloid leukemia cells inhibits proliferation and chromatid cohesion. Int J Mol Sci. 2020; 21:2073.9. Cui T, Bell EH, McElroy J, Liu K, Sebastian E, Johnson B, et al. A novel miR-146a-POU3F2/SMARCA5 pathway regulates stemness and therapeutic response in glioblastoma. Mol Cancer Res. 2021; 19:48–60.
Article10. Jin Q, Mao X, Li B, Guan S, Yao F, Jin F. Overexpression of SMARCA5 correlates with cell proliferation and migration in breast cancer. Tumour Biol. 2015; 36:1895–902.11. Ai C, Ma G, Deng Y, Zheng Q, Gen Y, Li W, et al. Nm23-H1 inhibits lung cancer bone-specific metastasis by upregulating miR-660-5p targeted SMARCA5. Thorac Cancer. 2020; 11:640–50.12. Gigek CO, Lisboa LC, Leal MF, Silva PN, Lima EM, Khayat AS, et al. SMARCA5 methylation and expression in gastric cancer. Cancer Invest. 2011; 29:162–6.13. Ding Y, Li Y, Zhao Z, Cliff Zhang Q, Liu F. The chromatin-remodeling enzyme Smarca5 regulates erythrocyte aggregation via Keap1-Nrf2 signaling. Elife. 2021; 10:e72557.14. Ding Y, Wang W, Ma D, Liang G, Kang Z, Xue Y, et al. Smarca5-mediated epigenetic programming facilitates fetal HSPC development in vertebrates. Blood. 2021; 137:190–202.
Article15. Dong YS, Hou WG, Li Y, Liu DB, Hao GZ, Zhang HF, et al. Unexpected requirement for a binding partner of the syntaxin family in phagocytosis by murine testicular Sertoli cells. Cell Death Differ. 2016; 23:787–800.16. Chai X, Yan J, Gao Y, Jin J. Endothelial HNF4α potentiates angiogenic dysfunction via enhancement of vascular endothelial growth factor resistance in T2DM. J Cell Biochem. 2019; 120:12989–3000.17. Hao JS, Zhu CJ, Yan BY, Yan CY, Ling R. Stimulation of KLF14/PLK1 pathway by thrombin signaling potentiates endothelial dysfunction in type 2 diabetes mellitus. Biomed Pharmacother. 2018; 99:859–66.18. He K, Qu H, Wang H, Zhang S, Qian XH, Li W. Regulated and functional expression of the corepressor MTA3 in rodent testis. Endocrinology. 2016; 157:4400–10.19. Fadini GP, Sartore S, Albiero M, Baesso I, Murphy E, Menegolo M, et al. Number and function of endothelial progenitor cells as a marker of severity for diabetic vasculopathy. Arterioscler Thromb Vasc Biol. 2006; 26:2140–6.20. Sawada N, Jiang A, Takizawa F, Safdar A, Manika A, Tesmenitsky Y, et al. Endothelial PGC-1α mediates vascular dysfunction in diabetes. Cell Metab. 2014; 19:246–58.21. Cheng S, Cui Y, Fan L, Mu X, Hua Y. T2DM inhibition of endothelial miR-342-3p facilitates angiogenic dysfunction via repression of FGF11 signaling. Biochem Biophys Res Commun. 2018; 503:71–8.22. Niccoli S, Boreham DR, Phenix CP, Lees SJ. Non-radioactive 2-deoxy-2-fluoro-D-glucose inhibits glucose uptake in xenograft tumours and sensitizes HeLa cells to doxorubicin in vitro. PLoS One. 2017; 12:e0187584.23. Chen YH, Lin SJ, Lin FY, Wu TC, Tsao CR, Huang PH, et al. High glucose impairs early and late endothelial progenitor cells by modifying nitric oxide-related but not oxidative stress-mediated mechanisms. Diabetes. 2007; 56:1559–68.24. Qi X, Okamoto Y, Murakawa T, Wang F, Oyama O, Ohkawa R, et al. Sustained delivery of sphingosine-1-phosphate using poly(lactic-co-glycolic acid)-based microparticles stimulates Akt/ERK-eNOS mediated angiogenesis and vascular maturation restoring blood flow in ischemic limbs of mice. Eur J Pharmacol. 2010; 634:121–31.25. Tie L, Li XJ, Wang X, Channon KM, Chen AF. Endothelium-specific GTP cyclohydrolase I overexpression accelerates refractory wound healing by suppressing oxidative stress in diabetes. Am J Physiol Endocrinol Metab. 2009; 296:E1423–9.26. Marshall CD, Hu MS, Leavitt T, Barnes LA, Cheung AT, Malhotra S, et al. Sanativo wound healing product does not accelerate reepithelialization in a mouse cutaneous wound healing model. Plast Reconstr Surg. 2017; 139:343–52.27. Fiorentino TV, Prioletta A, Zuo P, Folli F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr Pharm Des. 2013; 19:5695–703.28. Ning P, Zhong JG, Jiang F, Zhang Y, Zhao J, Tian F, et al. Role of protein S in castration-resistant prostate cancer-like cells. Endocr Relat Cancer. 2016; 23:595–607.29. Jin X, Zhang S, Ding T, Zhao P, Zhang C, Zhang Y, et al. Testicular Lmcd1 regulates phagocytosis by Sertoli cells through modulation of NFAT1/Txlna signaling pathway. Aging Cell. 2020; 19:e13217.30. Wang GG, Wang YZ, Xie J, Huang CY, Kong ZL, Ding X, et al. Cyclic tensile forces enhance the angiogenic properties of HUVECs by promoting the activities of human periodontal ligament cells. J Periodontol. 2021; 92:159–69.31. Luo Q, Wu X, Chang W, Zhao P, Zhu X, Chen H, et al. ARID1A hypermethylation disrupts transcriptional homeostasis to promote squamous cell carcinoma progression. Cancer Res. 2020; 80:406–17.32. Liu J, Li C, Wang J, Xu D, Wang H, Wang T, et al. Chromatin modifier MTA1 regulates mitotic transition and tumorigenesis by orchestrating mitotic mRNA processing. Nat Commun. 2020; 11:4455.33. Monteiro JP, Bennett M, Rodor J, Caudrillier A, Ulitsky I, Baker AH. Endothelial function and dysfunction in the cardiovascular system: the long non-coding road. Cardiovasc Res. 2019; 115:1692–704.34. Kumar S. SWI/SNF (BAF) complexes: from framework to a functional role in endothelial mechanotransduction. Curr Top Membr. 2021; 87:171–98.35. Griffin CT, Brennan J, Magnuson T. The chromatin-remodeling enzyme BRG1 plays an essential role in primitive erythropoiesis and vascular development. Development. 2008; 135:493–500.36. Huang X, Gao X, Diaz-Trelles R, Ruiz-Lozano P, Wang Z. Coronary development is regulated by ATP-dependent SWI/SNF chromatin remodeling component BAF180. Dev Biol. 2008; 319:258–66.37. Moonen JR, Chappell J, Shi M, Shinohara T, Li D, Mumbach MR, et al. KLF4 recruits SWI/SNF to increase chromatin accessibility and reprogram the endothelial enhancer landscape under laminar shear stress. Nat Commun. 2022; 13:4941.38. Ishikura K, Misu H, Kumazaki M, Takayama H, Matsuzawa-Nagata N, Tajima N, et al. Selenoprotein P as a diabetes-associated hepatokine that impairs angiogenesis by inducing VEGF resistance in vascular endothelial cells. Diabetologia. 2014; 57:1968–76.39. Alhusban A, Alkhazaleh E, El-Elimat T. Silymarin ameliorates diabetes-induced proangiogenic response in brain endothelial cells through a GSK-3β inhibition-induced reduction of VEGF release. J Diabetes Res. 2017; 2017:2537216.40. Xia L, Huang W, Bellani M, Seidman MM, Wu K, Fan D, et al. CHD4 has oncogenic functions in initiating and maintaining epigenetic suppression of multiple tumor suppressor genes. Cancer Cell. 2017; 31:653–68e7.41. McKenzie LD, LeClair JW, Miller KN, Strong AD, Chan HL, Oates EL, et al. CHD4 regulates the DNA damage response and RAD51 expression in glioblastoma. Sci Rep. 2019; 9:4444.42. Yun JH, Park SW, Kim KJ, Bae JS, Lee EH, Paek SH, et al. Endothelial STAT3 activation increases vascular leakage through downregulating tight junction proteins: implications for diabetic retinopathy. J Cell Physiol. 2017; 232:1123–34.43. Levings DC, Lacher SE, Palacios-Moreno J, Slattery M. Transcriptional reprogramming by oxidative stress occurs within a predefined chromatin accessibility landscape. Free Radic Biol Med. 2021; 171:319–31.44. Tommasi S, Pinto R, Danza K, Pilato B, Palumbo O, Micale L, et al. miR-151-5p, targeting chromatin remodeler SMARCA5, as a marker for the BRCAness phenotype. Oncotarget. 2016; 7:80363–72.45. Wu Z, Tang H, Xiong Q, Liu D, Xia T, Liang H, et al. Prognostic role of microRNA-205 in human gynecological cancer: a meta-analysis of fourteen studies. DNA Cell Biol. 2020; 39:875–89.46. Ouni M, Gottmann P, Westholm E, Schwerbel K, Jahnert M, Stadion M, et al. MiR-205 is up-regulated in islets of diabetes-susceptible mice and targets the diabetes gene Tcf7l2. Acta Physiol (Oxf). 2021; 232:e13693.47. Zhang Q, Xiao X, Li M, Li W, Yu M, Zhang H, et al. Acarbose reduces blood glucose by activating miR-10a-5p and miR-664 in diabetic rats. PLoS One. 2013; 8:e79697.48. Sugden WW, North TE. Making blood from the vessel: extrinsic and environmental cues guiding the endothelial-to-hematopoietic transition. Life (Basel). 2021; 11:1027.49. Kokavec J, Zikmund T, Savvulidi F, Kulvait V, Edelmann W, Skoultchi AI, et al. The ISWI ATPase Smarca5 (Snf2h) is required for proliferation and differentiation of hematopoietic stem and progenitor cells. Stem Cells. 2017; 35:1614–23.50. Norton KA, Popel AS. Effects of endothelial cell proliferation and migration rates in a computational model of sprouting angiogenesis. Sci Rep. 2016; 6:36992.51. Carmeliet P. VEGF as a key mediator of angiogenesis in cancer. Oncology. 2005; 69(Suppl 3):4–10.52. Oyama Y, Shigeta S, Tokunaga H, Tsuji K, Ishibashi M, Shibuya Y, et al. CHD4 regulates platinum sensitivity through MDR1 expression in ovarian cancer: a potential role of CHD4 inhibition as a combination therapy with platinum agents. PLoS One. 2021; 16:e0251079.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Effect of Acute Hyperglycemia on Endothelial Function in Type 2 Diabetes
- Hyperglycemia-induced Activation of Nuclear Transcription Factor kappaB in Cultured Fibroblasts and Endothelial Cells
- Effects of Acute Hyperglycemia on Endothelium-Dependent Vasodilation in Patients with Diabetes Mellitus or Impaired Glucose Metabolism
- Diabetes Mellitus and Endothelial Cell Dysfunction
- Effect of Diabetes in Surgery
