Enrichment of Wee1/CDC2 and NF-κB Signaling Pathway Constituents Mutually Contributes to CDDP Resistance in Human Osteosarcoma
- Affiliations
-
- 1Derpartment of Orthopedics, Shaoguan First People’s Hospital Affiliated to Southern Medical University, Guangdong, China
- 2Orthopedics Center, Zhujiang Hospital of Southern Medical University, Guangzhou, China
- 3Department of Orthopaedics, The Affiliated Yuebei People’s Hospital of Shantou University Medical College, Shaoguan, Guangdong, China
- 4The Second School of Clinical Medicine, Southern Medical University, Guangzhou, China
- 5Department of Pathology, Guangdong Provincal People’s Hospital & Guangdong, Academy of Medical Sciences, Guangzhou, China
- 6Guangzhou Laboratory (Guangzhou International Bio Island), Guangzhou, China
- 7University of Chinese Academy of Social Sciences (Graduate School), Guangzhou, China
- KMID: 2524609
- DOI: http://doi.org/10.4143/crt.2021.320
Abstract
- Purpose
Osteosarcoma (OS) universally exhibits heterogeneity and cisplatin (CDDP) resistance. Although the Wee1/CDC2 and nuclear factor кB (NF-κB) pathways were reported to show abnormal activation in some tumor cells with CDDP resistance, whether there is any concrete connection is currently unclear. We explored it in human OS cells.
Materials and Methods
Multiple OS cell lines were exposed to a Wee1 inhibitor (AZD1775) and CDDP to assess the half-maximal inhibitory concentration values. Western blot, coimmunoprecipitation, confocal immunofluorescence, cell cycle, and Cell Counting Kit-8assays were performed to explore the connection between the Wee1/CDC2 and NF-κB pathways and their subsequent physiological contribution to CDDP resistance. Finally, CDDP-resistant PDX-OS xenograft models were established to confirm that AZD1775 restores the antitumor effects of CDDP.
Results
A sensitivity hierarchy of OS cells to CDDP and AZD1775 exists. In the highly CDDP-tolerant cell lines, Wee1 and RelA were physically crosslinked, which resulted in increased abundance of phosphorylated CDC2 (Y15) and RelA (S536) and consequent modulation of cell cycle progression, survival, and proliferation. Wee1 inhibition restored the effects of CDDP on these processes in CDDP-resistant OS cells. In addition, animal experiments with CDDP-resistant PDX-OS cells showed that AZD1775 combined with CDDP not only restored CDDP efficacy but also amplified AZD1775 in inhibiting tumor growth and prolonged the median survival of the mice.
Conclusion
Simultaneous enrichment of molecules in the Wee1/CDC2 and NF-κB pathways and their consequent coactivation is a new molecular mechanism of CDDP resistance in OS cells. OS with this molecular signature may respond well to Wee1 inhibition as an alternative treatment strategy.
Keyword
Figure
-

Fig. 1 Wee1/CDC2 and nuclear factor κB (NF-κB) signaling pathway molecules in parental tumor cells had heterogeneous expression levels and were upregulated by cisplatin (CDDP). Tumor cells were seeded in a 10-cm cell culture dish (1×105 cells/dish) and cultured for 48 hours. Cells were treated with CDDP (25 μg/mL) in fresh medium for 6 hours when they reached 70%–80% confluence and then harvested. Parental cells were treated with fresh medium alone. Protein quantification was performed after cell lysis, and 30 μg protein per well was subjected to western blot analysis. The protein expression levels of Wee1/CDC2 and NF-κB pathway molecules IKKα, IKKβ, RelA, and IκB in conventional cells were significantly different, with the hierarchy Saos2 > U2OS > MG63 for OS cell lines (p < 0.01) (A), UN-SCC46 > UM-SCC47 > UM-SCC1 for head and neck squamous cell lines (p < 0.01) (B), and PDX-OS3 > PDX-OS2 > PDX-OS1 for PDX OS cell lines (p < 0.01) (C). CDDP significantly increased the protein expression levels of Wee1, IKKα, IKKβ, and RelA and decreased that of IκB in these tumor cells (p < 0.01).

Fig. 2 AZD1775 abolished the effects of tumor necrosis factor α (TNFα), cisplatin (CDDP), and Wee1 treatment in CDDP-tolerant osteosarcoma cells by amplifying the phosphorylation of both CDC2-Y15 and RelA-S536. Cells were seeded at 1×105 in a 10 cm cell culture dish, cultured for 48 hours, and harvested when the cells reached 70%–80% confluency. The cells were transfected with the plasmid Wee1 for 24 hours, after which they were treated with CDDP or TNFα for 6 hours. Vehicle treatment was used as a control. After cell lysis, the protein concentration was quantitated, and western blotting was performed with 25 μg of protein per sample. (A) In Saos2 cells, the levels of phosphorylated CDC2-Y15 (CDC2-Y15p) and RelA-S536 (RelA-S536p) in the TNFα treatment, Plasmid-Wee1 treatment and CDDP treatment groups were significantly amplified compared with that of the control group (p < 0.01) and were significantly reduced in the AZD1775 treatment group (p < 0.01). Compared with the TNFα treatment and Plasmid-Wee1 treatment groups, the Plasmid-Wee1+TNFα treatment group showed significantly increased CDC2-Y15p and RelA-S536p levels (p < 0.05). The levels of CDC2-Y15p and RelA-S536p in the TNFα+AZD1775 treatment group were significantly lower than those in the TNFα treatment group (p < 0.01), and the levels were significantly lower in the CDDP+AZD1775 treatment group than in the CDDP treatment group (p < 0.01). Moreover, these levels were significantly higher in the CDDP+TNFα treatment group than in the CDDP treatment group (p < 0.01). (B) In CDDP-resistant PDX-OS3 cells, the levels of CDC2-Y15 and RelA-S536 in each treatment group were basically consistent with those in the corresponding Saos2 cells. Interestingly, CDC2-Y15p and RelA-S536p levels were more obviously decreased in the treatment groups containing AZD1775, with statistical significance (p < 0.01). (C) Although the levels of CDC2-Y15p and RelA-S536 in PDX-OS1 cells were significantly increased in the CDDP-treated group compared to the control group, the levels of CDC2-Y15p in the CDDP+AZD1775 treatment group were decreased (p < 0.01), which was accompanied by increased RelA-S536 phosphorylation. In addition, the levels of RelA-S536p were amplified in the CDDP+TNFα group (p < 0.01), accompanied by a reduction in CDC2-Y15p levels. This is consistent with the results in U2OS and MG63 cells (data not shown).

Fig. 3 The interaction between Wee1 and RelA, as indicated by coimmunocipitation assays in osteosarcoma cells. Cells were seeded at 1×105 in a 10 cm cell culture dish and cultured for 48 hours. Cells were treated with different concentrations of cisplatin (CDDP; 5, 15, and 45 μg/mL) for 6 hours when they reached 70%–80% confluence and then harvested. Whole-cell lysate was used as the positive control, and IgG was used as the negative control. Protein quantification was performed after cell lysis, and the corresponding immunoprecipitants were obtained according to the method instructions, with 30 μg of immunoprecipitants per sample subjected to western blotting. In Saos2 cells (A), Wee1 binding to RelA showed a CDDP dose-dependent effect when treated with three different concentrations of CDDP. This result was reproduced in PDX-OS3 (B) and U2OS cells (C).

Fig. 4 Wee1 affects the nuclear translocation of RelA in cisplatin (CDDP)-resistant osteosarcoma cells. In confocal immunofluorescence experiments, PDX-OS3 cells were seeded in a cell culture plate at 2×104, cultured for 48 hours, treated with CDDP (25 μg/mL) for 6 hours when the cells reached 50%–60% confluence, and harvested. DMSO was used as a negative control. In PDX-OS3 cells, both Wee1 (green) and RelA (red) fluorescence signals were enhanced in the CDDP treatment group compared to the control group (row 1). Compared with the CDDP treatment group (row 2), the CDDP+AZD1775 group (row 3) showed Wee1 green signal distribution and aggregation mainly in the cytoplasm, while the RelA red signals were congested in the periphery of the nucleus and surrounded by Wee1 green signals. Furthermore, the RelA red signal in the nucleus was reduced. In addition, compared to the CDDP group, the CDDP+si-Wee1 treatment group (row 4) showed that the Wee1 green signal was obviously reduced, while the RelA red signal was also congested around the nucleus and was significantly reduced in the nucleus, which is consistent with the results from the CDDP+AZD1775 group. Scale bars=20 μm.
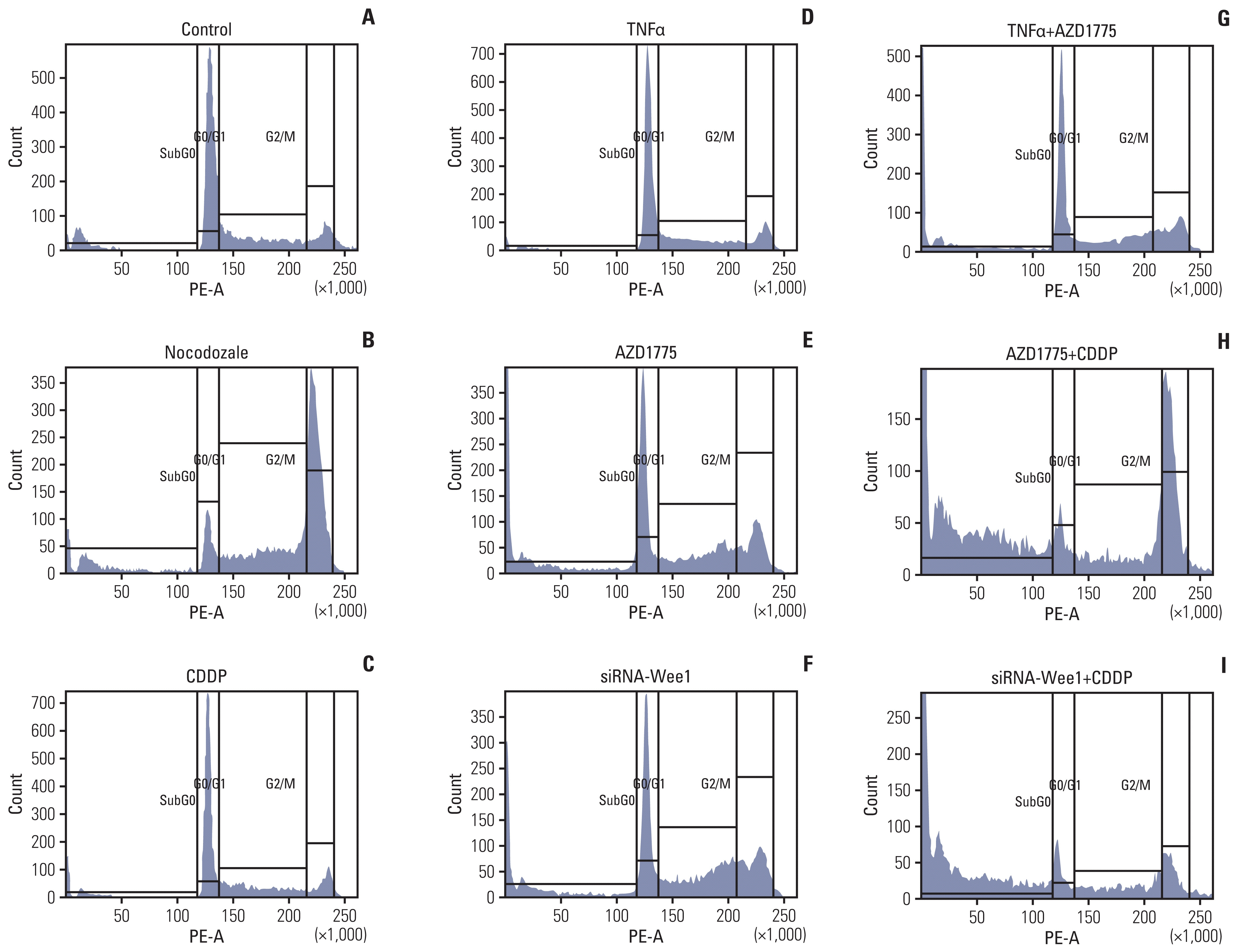
Fig. 5 Inhibition of Wee1 restores the effects of cisplatin (CDDP) on cell cycle progression and survival in Saos2 cells. In the cell cycle assays, Saos2 cells were seeded in a 10 cm cell culture dish at 1×105 and cultured for 48 hours. Cells were treated with si-Wee1 transfection or CDDP, nocodozale, tumor necrosis factor α (TNFα) and other treatments for 24 hours when the cells reached 70%–80% confluence, after which they were harvested. Vehicle treatment was used as a negative control (A), and the cell cycle G2 inhibitor nocodozale was used as a positive control (B). The proportion of G1 phase cells in the nocodozale treatment group was significantly decreased, while the proportion of G2 phase cells was significantly increased (p < 0.01). Compared with the negative control group, there was no significant difference in the proportion of cells in the different phases in the CDDP (C) or TNFα groups (D). Compared with the negative control and positive control groups, the proportion of cells in G2 and sub-G0 phases (cells dying after mitotic catastrophes) and the proportion of cells in G1 phase in the AZD1775 group (E) and siRNA-Wee1 group (F) were significantly increased, whereas the proportion of G1 phase cells was significantly decreased (p < 0.01). Compared with the TNFα treatment group (D), the AZD1775+TNFα treatment group (G) showed an increased proportion of G2 phase cells but a significant decrease in the proportion of sub-G0 phase cells (p < 0.01). Compared with the CDDP treatment group, the AZD1775+CDDP group (H) and siRNA-Wee1+CDDP group (I) showed significant decreases in the proportion of G1 phase cells (p < 0.01) and significant increases in the proportion of sub-G0 cell fragments (p < 0.01).

Fig. 6 Inhibition of Wee1 restores the effects of cisplatin (CDDP) on cell proliferation and activation of cell death-related molecular pathways in osteosarcoma cells with high tolerance to CDDP. In the Saos2 cell proliferation experiment (A), there was no significant difference in the cell optical density (OD) value between the tumor necrosis factor α (TNFα) and CDDP treatment groups and the control group. Compared with the control and CDDP treatment groups, the AZD1775 treatment and siRNA-Wee1 treatment groups showed significantly smaller OD values (p < 0.05). Compared with the TNFα group, the AZD1775+TNFα group also showed significantly lower OD values (p < 0.05). Compared with the CDDP treatment group, the CDDP+AZD1775 treatment group and CDDP+si-Wee1 treatment group both had the most significant decreases in OD values (p < 0.01). In the PDX-OS3 cell proliferation experiment (B), similar to those in Saos2 cells, the OD value was significantly lower in the AZD1775 and siRNA-Wee1 treatment groups than in the control group (p < 0.05) and was significantly lower in the AZD1775+TNFα treatment group than in the TNFα treatment group (p < 0.05). Compared with the CDDP treatment group, the CDDP+AZD1775 treatment group and CDDP+si-Wee1 treatment group both had the most significant decreases in the OD value (p < 0.01). In the Saos2 western blot assay (C), compared with the control group, the CDDP treatment group showed a significant increase in the levels of the NF-κB downstream molecules Bcl-2 and CCND1 (p < 0.05), while the differences in the expression of the apoptosis-related proteins cleaved poly(ADP-ribose) polymerase (PARP) and cleaved caspase-3 were not statistically significant. Compared with the control group, the AZD1775 or siRNA-Wee1 treatment group showed significantly lower Bcl-2 and CCND1 expression levels (p < 0.05) and significantly higher cleaved PARP and cleaved caspase-3 levels (p < 0.05). Moreover, Bcl-2 and CCND1 levels were significantly decreased in the CDDP+AZD1775 group and CDDP+si-Wee1 group (p < 0.01), which were accompanied by simultaneous increases in the apoptosis-related proteins cleaved PARP and cleaved caspase-3 (p < 0.01). In PDX-OS3 cells (D), consistent with the results in Saos2 cells, Wee1 inhibition treatment significantly decreased Bcl-2 and CCND1 expression and simultaneously increased cleaved PARP and cleaved caspase-3 levels, especially in the CDDP combined with Wee1 inhibition group.

Fig. 7 AZD1775 combined with cisplatin (CDDP) treatment enhanced and restored the therapeutic effect of CDDP, resulting in an extended median survival of the host. PDX-OS3 cells were transplanted into female nude mice and grown for 4 weeks, and the mice were randomly divided into four groups (5 mice per group) (A-D). CDDP 5 mg/kg was intraperitoneally injected once a week, and AZD1775 50 mg/kg was intragastrically administered twice a week for 3 weeks (E). Tumor volume changes were recorded regularly (F), and survival analysis was performed (G). Compared with the control group (A), the CDDP treatment group (B) showed a slightly decreased tumor volume, but the difference was not significant (p > 0.05). However, the tumor volume was significantly decreased in the CDDP+AZD775 group (C) (p < 0.01). The tumor volume was also significantly reduced in the AZD1775 treatment alone group (D) compared with the control group or CDDP treatment group (p < 0.05). The median survival of mice in the control, CDDP treatment, AZD1775 treatment and CDDP+AZD1775 treatment groups was 51, 53, 72, and 89 days, respectively. The median survival of the CDDP+AZD1775 treatment group was the longest, and the difference was significant (p < 0.01).

Fig. 8 Molecular mechanism of the synergistic interaction of Wee1 with RelA to promote cisplatin (CDDP). NF-κB, nuclear factor κB; TNFα, tumor necrosis factor α.
Reference
-
References
1. Isakoff MS, Bielack SS, Meltzer P, Gorlick R. Osteosarcoma: current treatment and a collaborative pathway to success. J Clin Oncol. 2015; 33:3029–35.
Article2. Makovec T. Cisplatin and beyond: molecular mechanisms of action and drug resistance development in cancer chemotherapy. Radiol Oncol. 2019; 53:148–58.
Article3. Dagogo-Jack I, Shaw AT. Tumour heterogeneity and resistance to cancer therapies. Nat Rev Clin Oncol. 2018; 15:81–94.
Article4. Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol. 2014; 740:364–78.
Article5. Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol. 2005; 5:749–59.6. Wang Y, Fu Q, Zhao W. Tetramethylpyrazine inhibits osteosarcoma cell proliferation via downregulation of NF-kappaB in vitro and in vivo. Mol Med Rep. 2013; 8:984–8.7. Gong Y, Zack TI, Morris LG, Lin K, Hukkelhoven E, Raheja R, et al. Pan-cancer genetic analysis identifies PARK2 as a master regulator of G1/S cyclins. Nat Genet. 2014; 46:588–94.
Article8. Miranda MA, Marcato PD, Mondal A, Chowdhury N, Gebeyehu A, Surapaneni SK, et al. Cytotoxic and chemosensitizing effects of glycoalkaloidic extract on 2D and 3D models using RT4 and patient derived xenografts bladder cancer cells. Mater Sci Eng C Mater Biol Appl. 2021; 119:111460.
Article9. Conde de la Rosa L, Schoemaker MH, Vrenken TE, Buist-Homan M, Havinga R, Jansen PL, et al. Superoxide anions and hydrogen peroxide induce hepatocyte death by different mechanisms: involvement of JNK and ERK MAP kinases. J Hepatol. 2006; 44:918–29.
Article10. Nam AR, Jin MH, Bang JH, Oh KS, Seo HR, Oh DY, et al. Inhibition of ATR increases the sensitivity to WEE1 inhibitor in biliary tract cancer. Cancer Res Treat. 2020; 52:945–56.
Article11. Yan M, Ni J, Song D, Ding M, Huang J. Activation of unfolded protein response protects osteosarcoma cells from cisplatin-induced apoptosis through NF-kappaB pathway. Int J Clin Exp Pathol. 2015; 8:10204–15.12. Moiseeva TN, Qian C, Sugitani N, Osmanbeyoglu HU, Bakkenist CJ. WEE1 kinase inhibitor AZD1775 induces CDK1 kinase-dependent origin firing in unperturbed G1- and S-phase cells. Proc Natl Acad Sci U S A. 2019; 116:23891–3.
Article13. Geenen JJ, Schellens JH. Molecular pathways: targeting the protein kinase Wee1 in cancer. Clin Cancer Res. 2017; 23:4540–4.
Article14. Kuijjer ML, Hogendoorn PC, Cleton-Jansen AM. Genome-wide analyses on high-grade osteosarcoma: making sense of a genomically most unstable tumor. Int J Cancer. 2013; 133:2512–21.
Article15. Castillo-Tandazo W, Mutsaers AJ, Walkley CR. Osteosarcoma in the post genome era: preclinical models and approaches to identify tractable therapeutic targets. Curr Osteoporos Rep. 2019; 17:343–52.
Article16. Lu W, Chao T, Ruiqi C, Juan S, Zhihong L. Patient-derived xenograft models in musculoskeletal malignancies. J Transl Med. 2018; 16:107.
Article17. Li W, Zhang X, Xi X, Li Y, Quan H, Liu S, et al. PLK2 modulation of enriched TAp73 affects osteogenic differentiation and prognosis in human osteosarcoma. Cancer Med. 2020; 9:4371–85.
Article18. Lu H, Yang X, Duggal P, Allen CT, Yan B, Cohen J, et al. TNF-alpha promotes c-REL/DeltaNp63alpha interaction and TAp73 dissociation from key genes that mediate growth arrest and apoptosis in head and neck cancer. Cancer Res. 2011; 71:6867–77.19. Blattmann C, Thiemann M, Stenzinger A, Roth EK, Dittmar A, Witt H, et al. Establishment of a patient-derived orthotopic osteosarcoma mouse model. J Transl Med. 2015; 13:136.
Article20. Hu ZB, Liao XH, Xu ZY, Yang X, Dong C, Jin AM, et al. PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells. Cancer Med. 2016; 5:74–87.21. Cho JG, Choi JS, Lee JH, Cho MG, Kim HS, Noh HD, et al. MED28 over-expression shortens the cell cycle and induces genomic instability. Int J Mol Sci. 2019; 20:1746.
Article22. Duan Z, Gao Y, Shen J, Choy E, Cote G, Harmon D, et al. miR-15b modulates multidrug resistance in human osteosarcoma in vitro and in vivo. Mol Oncol. 2017; 11:151–66.23. Lin X, Chen D, Zhang C, Zhang X, Li Z, Dong B, et al. Augmented antitumor activity by olaparib plus AZD1775 in gastric cancer through disrupting DNA damage repair pathways and DNA damage checkpoint. J Exp Clin Cancer Res. 2018; 37:129.
Article24. Woo XY, Giordano J, Srivastava A, Zhao ZM, Lloyd MW, de Bruijn R, et al. Conservation of copy number profiles during engraftment and passaging of patient-derived cancer xenografts. Nat Genet. 2021; 53:86–99.25. Teng S, Li YE, Yang M, Qi R, Huang Y, Wang Q, et al. Tissue-specific transcription reprogramming promotes liver metastasis of colorectal cancer. Cell Res. 2020; 30:34–49.
Article26. Stewart E, Federico SM, Chen X, Shelat AA, Bradley C, Gordon B, et al. Orthotopic patient-derived xenografts of paediatric solid tumours. Nature. 2017; 549:96–100.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Eriodictyol Inhibits the Production and Gene Expression of MUC5AC Mucin via the IκBα-NF-κB p65 Signaling Pathway in Airway Epithelial Cells
- Wnt-C59 inhibits proinflammatory cytokine expression by reducing the interaction between β-catenin and NF-κB in LPS-stimulated epithelial and macrophage cells
- Porcine parvovirus nonstructural protein NS1 activates NF-κB and it involves TLR2 signaling pathway
- Activation of CpG-ODN-Induced TLR9 Signaling Inhibited by Interleukin-37 in U937 Human Macrophages
- Kaempferol Regulates the Expression of Airway MUC5AC Mucin Gene via IκBα-NF-κB p65 and p38-p44/42-Sp1 Signaling Pathways
