Effects of Lipopolysaccharide on Oligodendrocyte Differentiation at Different Developmental Stages: an In Vitro Study
- Affiliations
-
- 1Department of Pediatrics, Hanyang University College of Medicine, Seoul, Korea
- 2Kangwon National University College of Veterinary Medicine, Chuncheon, Korea
- 3Department of Pediatrics, Hanyang University Guri Hospital, Guri, Korea
- 4Department of Child Psychotherapy, Hanyang University Graduate School of Medicine, Seoul, Korea
- 5Department of Pediatrics, Seoul University College of Medicine, Seoul, Korea
- KMID: 2523552
- DOI: http://doi.org/10.3346/jkms.2021.36.e332
Abstract
- Background
Lipopolysaccharide (LPS) exerts cytotoxic effects on brain cells, especially on those belonging to the oligodendrocyte lineage, in preterm infants. The susceptibility of oligodendrocyte lineage cells to LPS-induced inflammation is dependent on the developmental stage. This study aimed to investigate the effect of LPS on oligodendrocyte lineage cells at different developmental stages in a microglial cell and oligodendrocyte coculture model.
Methods
The primary cultures of oligodendrocytes and microglia cells were prepared from the forebrains of 2-day-old Sprague–Dawley rats. The oligodendrocyte progenitor cells (OPCs) co-cultured with microglial cells were treated with 0 (control), 0.01, 0.1, and 1 µg/mL LPS at the D3 stage to determine the dose of LPS that impairs oligodendrocyte differentiation. The co-culture was treated with 0.01 µg/mL LPS, which was the lowest dose that did not impair oligodendrocyte differentiation, at the developmental stages D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group). On day 7 of differentiation, oligodendrocytes were subjected to neural glial antigen 2 (NG2) and myelin basic protein (MBP) immunostaining to examine the number of OPCs and mature oligodendrocytes, respectively.
Results
LPS dose-dependently decreased the proportion of mature oligodendrocytes (MBP+ cells) relative to the total number of cells. The number of MBP+ cells in the early LPS group was significantly lower than that in the late LPS group. Compared with those in the control group, the MBP+ cell numbers were significantly lower and the NG2+ cell numbers were significantly higher in the double LPS group, which exhibited impaired oligodendrocyte lineage cell development, on day 7 of differentiation.
Conclusion
Repetitive LPS stimulation during development significantly inhibited brain cell development by impairing oligodendrocyte differentiation. In contrast, brain cell development was not affected in the late LPS group. These findings suggest that inflammation at the early developmental stage of oligodendrocytes increases the susceptibility of the preterm brain to inflammation-induced injury.
Figure
-
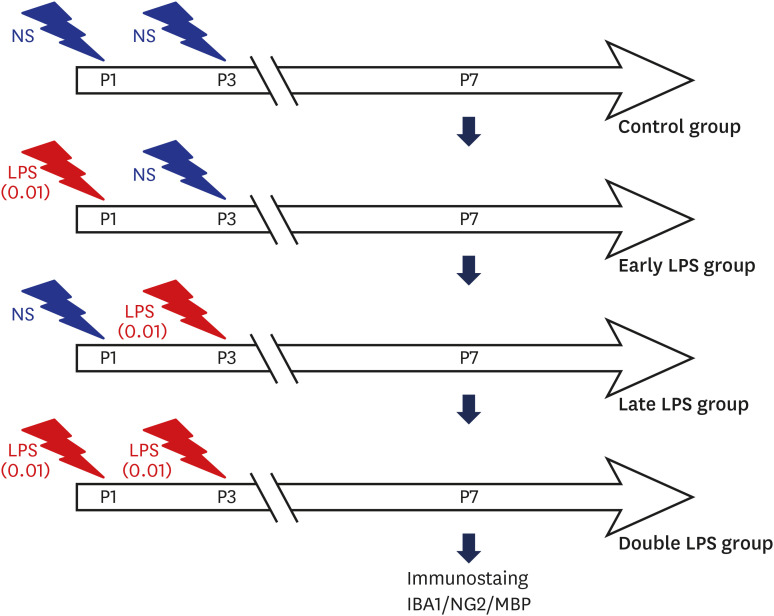
Fig. 1 Schematic outline of the experimental protocol. The oligodendrocyte-microglial cell co-culture was treated with the lowest dose of LPS (0.01 µg/mL), which does not inhibit oligodendrocyte differentiation, at the developmental stages D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group).LPS = lipopolysaccharide, MBP = myelin basic protein.
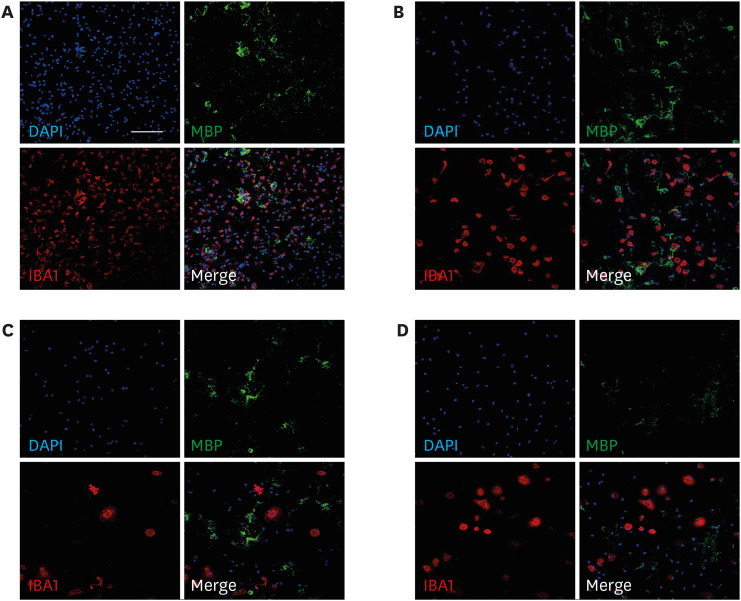
Fig. 2 Effect of various doses of LPS on oligodendrocyte differentiation. On day 3, the co-culture was treated with 0.01, 0.1, and 1 µg/mL LPS. On day 7 of differentiation, the nuclei were counterstained with 4′,6-diamidino-2-phenylindole (blue). The oligodendrocyte-microglia co-culture was subjected to MBP (green; mature oligodendrocyte marker) and IBA1 (red; microglia marker) immunostaining. The co-culture was treated with vehicle (control) (A) and 0.01 (B), 0.1 (C), and 1 µg/mL LPS (D). Scale bar = 100 µm (200 × magnification).LPS = lipopolysaccharide, MBP = myelin basic protein.
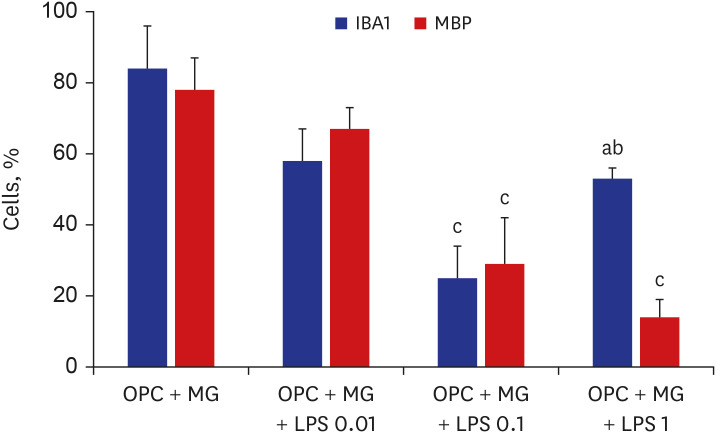
Fig. 3 Representative images of oligodendrocytes treated with various doses of LPS on day 7 of differentiation (n = 4/group).LPS = lipopolysaccharide, OPC = oligodendrocyte progenitor cell, MBP = myelin basic protein, MG = microglia.P values from Mann-Whitney U test: a0.007 vs. control; b0.009 vs. OPC + MG + LPS 0.1. P values from one-way analysis of variance analysis of variance with Bonferroni: c< 0.001 vs. control.
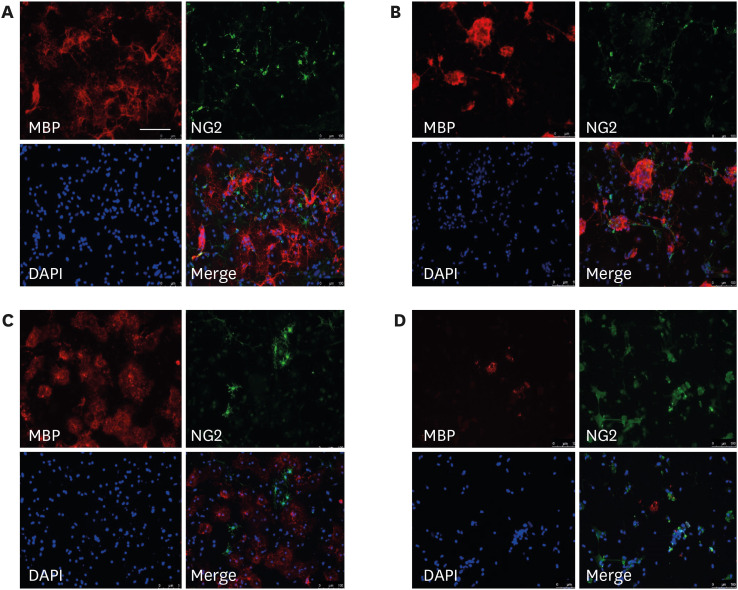
Fig. 4 Representative images of oligodendrocyte-microglia co-culture following different schedules of LPS treatment. Representative images of the oligodendrocyte-microglia co-culture subjected to immunostaining after in vitro treatment with vehicle (control) (A) or 0.01 µg/mL LPS at D1 (early LPS group) (B), D3 (late LPS group) (C), or D1 and D3 (double LPS group) stages. Scale bar = 100 µm (200 × magnification).LPS = lipopolysaccharide, MBP = myelin basic protein.
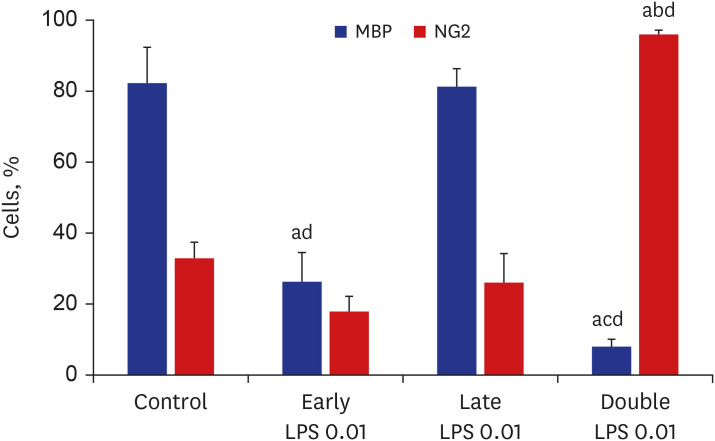
Fig. 5 Quantification of the relative numbers of NG2+ and MBP+ cells. Relative numbers of NG2+ and MBP+ cells were quantified after treatment with vehicle (control) or 0.01 µg/mL LPS at D1 (early LPS group), D3 (late LPS group), or D1 and D3 (double LPS group) stages. The number of MBP+ cells in the early LPS group was significantly lower than that in the control group. Compared with those in the control group, the MBP+ cell numbers were significantly lower and the NG2+ cell numbers were significantly higher in the double LPS group at day 7 of differentiation (n = 5/group).LPS = lipopolysaccharide, MBP = myelin basic protein.All P values from one-way analysis of variance analysis of variance with Bonferroni; a< 0.001 vs. control; b< 0.001 vs. early LPS; c0.004 vs. early LPS; d< 0.001 vs. late LPS.
Reference
-
1. Vieira ME, Linhares MB. Quality of life of individuals born preterm: a systematic review of assessment approaches. Qual Life Res. 2016; 25(9):2123–2139. PMID: 26995563.
Article2. Volpe JJ. Neurobiology of periventricular leukomalacia in the premature infant. Pediatr Res. 2001; 50(5):553–562. PMID: 11641446.
Article3. Back SA, Rivkees SA. Emerging concepts in periventricular white matter injury. Semin Perinatol. 2004; 28(6):405–414. PMID: 15693397.
Article4. Back SA. Cerebral white and gray matter injury in newborns: new insights into pathophysiology and management. Clin Perinatol. 2014; 41(1):1–24. PMID: 24524444.5. Wang X, Rousset CI, Hagberg H, Mallard C. Lipopolysaccharide-induced inflammation and perinatal brain injury. Semin Fetal Neonatal Med. 2006; 11(5):343–353. PMID: 16793357.
Article6. Sherwin C, Fern R. Acute lipopolysaccharide-mediated injury in neonatal white matter glia: role of TNF-alpha, IL-1beta, and calcium. J Immunol. 2005; 175(1):155–161. PMID: 15972642.7. Pang Y, Campbell L, Zheng B, Fan L, Cai Z, Rhodes P. Lipopolysaccharide-activated microglia induce death of oligodendrocyte progenitor cells and impede their development. Neuroscience. 2010; 166(2):464–475. PMID: 20035837.
Article8. Back SA, Gan X, Li Y, Rosenberg PA, Volpe JJ. Maturation-dependent vulnerability of oligodendrocytes to oxidative stress-induced death caused by glutathione depletion. J Neurosci. 1998; 18(16):6241–6253. PMID: 9698317.
Article9. Back SA, Riddle A, McClure MM. Maturation-dependent vulnerability of perinatal white matter in premature birth. Stroke. 2007; 38(2):Suppl. 724–730. PMID: 17261726.
Article10. Back SA, Han BH, Luo NL, Chricton CA, Xanthoudakis S, Tam J, et al. Selective vulnerability of late oligodendrocyte progenitors to hypoxia-ischemia. J Neurosci. 2002; 22(2):455–463. PMID: 11784790.
Article11. Miller BA, Crum JM, Tovar CA, Ferguson AR, Bresnahan JC, Beattie MS. Developmental stage of oligodendrocytes determines their response to activated microglia in vitro. J Neuroinflammation. 2007; 4(1):28. PMID: 18039385.
Article12. Shah DK, Doyle LW, Anderson PJ, Bear M, Daley AJ, Hunt RW, et al. Adverse neurodevelopment in preterm infants with postnatal sepsis or necrotizing enterocolitis is mediated by white matter abnormalities on magnetic resonance imaging at term. J Pediatr. 2008; 153(2):170–175. –175.e1. PMID: 18534228.
Article13. Glass TJ, Chau V, Grunau RE, Synnes A, Guo T, Duerden EG, et al. Multiple postnatal infections in newborns born preterm predict delayed maturation of motor pathways at term-equivalent age with poorer motor outcomes at 3 years. J Pediatr. 2018; 196:91–97.e1. PMID: 29398063.
Article14. Lee I, Neil JJ, Huettner PC, Smyser CD, Rogers CE, Shimony JS, et al. The impact of prenatal and neonatal infection on neurodevelopmental outcomes in very preterm infants. J Perinatol. 2014; 34(10):741–747. PMID: 25033076.
Article15. Kim HS, Kim EK, Park HK, Ahn DH, Kim MJ, Lee HJ. Cognitive outcomes of children with very low birth weight at 3 to 5 years of age. J Korean Med Sci. 2020; 35(1):e4. PMID: 31898433.
Article16. Shim SY, Cho SJ, Park EA. Neurodevelopmental outcomes at 18-24 months of corrected age in very low birth weight infants with late-onset sepsis. J Korean Med Sci. 2021; 36(35):e205. PMID: 34490752.
Article17. Galinsky R, Lear CA, Dean JM, Wassink G, Dhillon SK, Fraser M, et al. Complex interactions between hypoxia-ischemia and inflammation in preterm brain injury. Dev Med Child Neurol. 2018; 60(2):126–133. PMID: 29194585.
Article18. Markus T, Cronberg T, Cilio C, Pronk C, Wieloch T, Ley D. Tumor necrosis factor receptor-1 is essential for LPS-induced sensitization and tolerance to oxygen-glucose deprivation in murine neonatal organotypic hippocampal slices. J Cereb Blood Flow Metab. 2009; 29(1):73–86. PMID: 18728678.
Article19. Lehnardt S, Lachance C, Patrizi S, Lefebvre S, Follett PL, Jensen FE, et al. The toll-like receptor TLR4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J Neurosci. 2002; 22(7):2478–2486. PMID: 11923412.
Article20. Hoogland IC, Houbolt C, van Westerloo DJ, van Gool WA, van de Beek D. Systemic inflammation and microglial activation: systematic review of animal experiments. J Neuroinflammation. 2015; 12(1):114. PMID: 26048578.
Article21. Liu B, Wang K, Gao HM, Mandavilli B, Wang JY, Hong JS. Molecular consequences of activated microglia in the brain: overactivation induces apoptosis. J Neurochem. 2001; 77(1):182–189. PMID: 11279274.
Article22. Liu B, Du L, Hong JS. Naloxone protects rat dopaminergic neurons against inflammatory damage through inhibition of microglia activation and superoxide generation. J Pharmacol Exp Ther. 2000; 293(2):607–617. PMID: 10773035.23. Kim WG, Mohney RP, Wilson B, Jeohn GH, Liu B, Hong JS. Regional difference in susceptibility to lipopolysaccharide-induced neurotoxicity in the rat brain: role of microglia. J Neurosci. 2000; 20(16):6309–6316. PMID: 10934283.
Article24. Chang RC, Chen W, Hudson P, Wilson B, Han DS, Hong JS. Neurons reduce glial responses to lipopolysaccharide (LPS) and prevent injury of microglial cells from over-activation by LPS. J Neurochem. 2001; 76(4):1042–1049. PMID: 11181823.
Article25. Semple BD, Blomgren K, Gimlin K, Ferriero DM, Noble-Haeusslein LJ. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog Neurobiol. 2013; 106-107:1–16. PMID: 23583307.
Article26. Glass HC, Bonifacio SL, Chau V, Glidden D, Poskitt K, Barkovich AJ, et al. Recurrent postnatal infections are associated with progressive white matter injury in premature infants. Pediatrics. 2008; 122(2):299–305. PMID: 18676547.
Article27. Korzeniewski SJ, Romero R, Cortez J, Pappas A, Schwartz AG, Kim CJ, et al. A “multi-hit” model of neonatal white matter injury: cumulative contributions of chronic placental inflammation, acute fetal inflammation and postnatal inflammatory events. J Perinat Med. 2014; 42(6):731–743. PMID: 25205706.
Article28. Hagberg H, Gressens P, Mallard C. Inflammation during fetal and neonatal life: implications for neurologic and neuropsychiatric disease in children and adults. Ann Neurol. 2012; 71(4):444–457. PMID: 22334391.
Article29. Cornette L. Fetal and neonatal inflammatory response and adverse outcome. Semin Fetal Neonatal Med. 2004; 9(6):459–470. PMID: 15691784.
Article30. Dammann O, Kuban KC, Leviton A. Perinatal infection, fetal inflammatory response, white matter damage, and cognitive limitations in children born preterm. Ment Retard Dev Disabil Res Rev. 2002; 8(1):46–50. PMID: 11921386.
Article31. Malaeb S, Dammann O. Fetal inflammatory response and brain injury in the preterm newborn. J Child Neurol. 2009; 24(9):1119–1126. PMID: 19605775.
Article32. Segovia KN, McClure M, Moravec M, Luo NL, Wan Y, Gong X, et al. Arrested oligodendrocyte lineage maturation in chronic perinatal white matter injury. Ann Neurol. 2008; 63(4):520–530. PMID: 18393269.
Article33. Billiards SS, Haynes RL, Folkerth RD, Borenstein NS, Trachtenberg FL, Rowitch DH, et al. Myelin abnormalities without oligodendrocyte loss in periventricular leukomalacia. Brain Pathol. 2008; 18(2):153–163. PMID: 18177464.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Microarray Screening for Genes Involved in Oligodendrocyte Differentiation in the Zebrafish CNS
- Estimation of Mandibular Third Molar Development Using the Correlation in Dental Developmental Stages
- Assessment of Developmental Toxicants using Human Embryonic Stem Cells
- Identification of Long Non-Coding RNAs and Their Target Genes from Mycelium and Primordium in Model Mushroom Schizophyllum commune
- In vitro Model to Assess Neurotoxicity Induced by Anticancer Chemotherapeutic Agents
