Vimentin Deficiency Prevents High-Fat Diet-Induced Obesity and Insulin Resistance in Mice
- Affiliations
-
- 1Department of Molecular Medicine, College of Medicine, Ewha Womans University, Seoul, Korea.
- 2Immune and Vascular Cell Network Research Center, National Creative Initiatives, Department of Life Sciences, Ewha Womans University, Seoul, Korea.
- KMID: 2514078
- DOI: http://doi.org/10.4093/dmj.2019.0198
Abstract
Background Obesity and type 2 diabetes mellitus are world-wide health problems, and lack of understanding of their linking mechanism is one reason for limited treatment options. We determined if genetic deletion of vimentin, a type 3 intermediate filament, affects obesity and type 2 diabetes mellitus.
Methods We fed vimentin-null (
Vim −/−) mice and wild-type mice a high-fat diet (HFD) for 10 weeks and measured weight change, adiposity, blood lipids, and glucose. We performed intraperitoneal glucose tolerance tests and measured CD36, a major fatty acid translocase, and glucose transporter type 4 (GLUT4) in adipocytes from both groups of mice.Results Vim −/− mice fed an HFD showed less weight gain, less adiposity, improved glucose tolerance, and lower serum level of fasting glucose. However, serum triglyceride and non-esterified fatty acid levels were higher inVim −/− mice than in wild-type mice. Vimentin-null adipocytes showed 41.1% less CD36 on plasma membranes, 27% less uptake of fatty acids, and 50.3% less GLUT4, suggesting defects in intracellular trafficking of these molecules.Conclusion We concluded that vimentin deficiency prevents obesity and insulin resistance in mice fed an HFD and suggest vimentin as a central mediator linking obesity and type 2 diabetes mellitus.
Figure
-
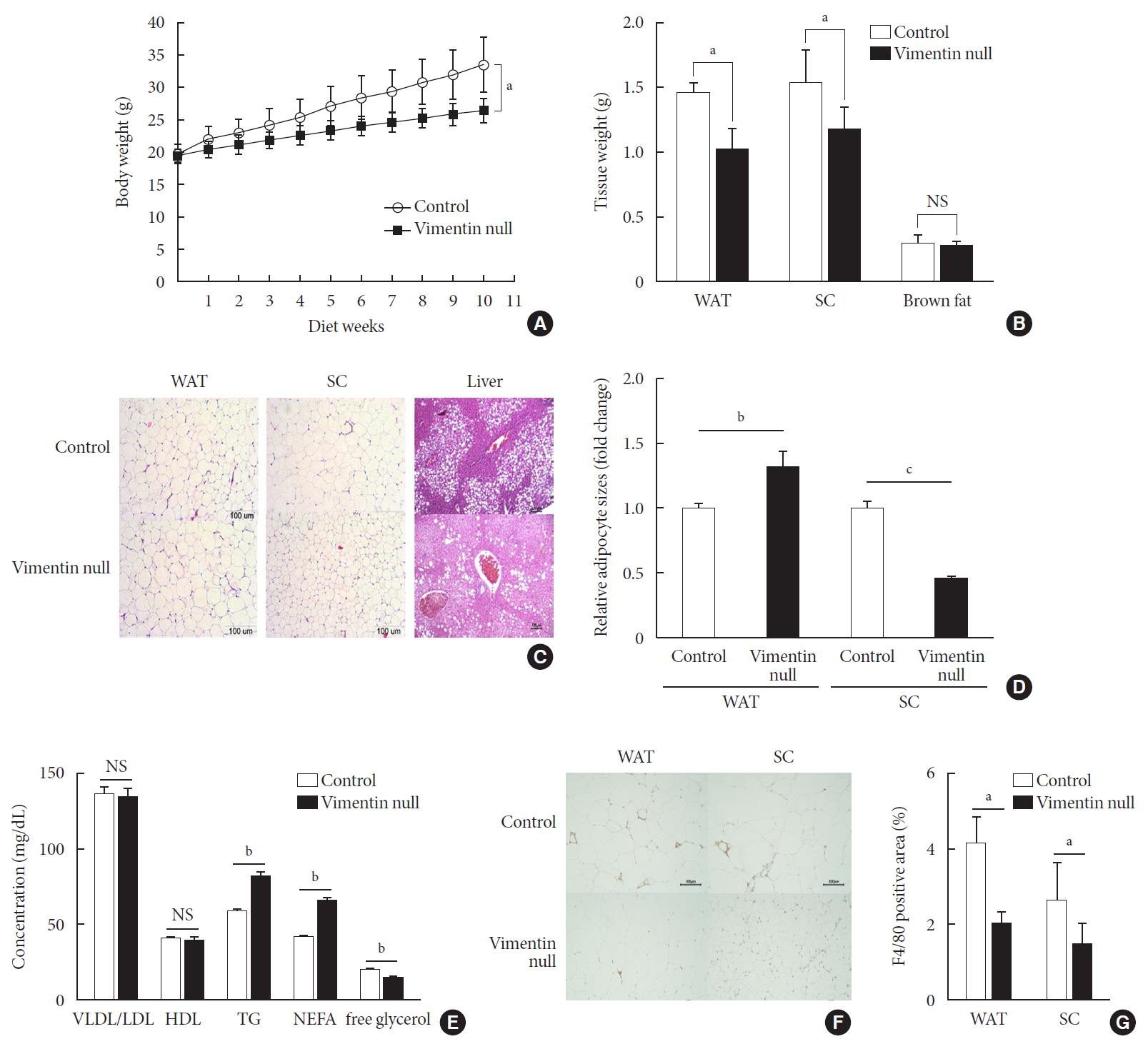
Fig. 1 Vimentin deficiency prevents diet-induced obesity. (A) Vim−/− mice (n=10) and control wild-type mice (n=10) were fed a high-fat diet (HFD) for 10 weeks. Body weight changes of control and Vim−/− mice on an HFD. Bars indicate mean±standard error of the mean (SEM). (B) Weights of adipose tissues from the experimental mice in (A). Bars indicate mean±SEM. (C) Representative images of adipose tissues and livers harvested from the experimental mice in (A) with hematoxylin-eosin staining. Scale bar: 100 µm. (D) The graph shows comparisons of adipocyte sizes between Vim−/− mice and wide-type mice. Adipocytes from three tissue sections per mouse type were measured. Each bar denotes the mean±SEM. (E) Concentrations of very low density lipoprotein/low density lipoprotein (VLDL/LDL), high density lipoprotein (HDL), triglyceride (TG), non-esterified fatty acid (NEFA), and free glycerol in the sera of the Vim−/− and control mice in Fig. 1A. The graph shows mean±SEM. (F) Representative images of adipose tissues from the experimental mice in (A), stained with antibody for the macrophage marker F4/80. Scale bar: 100 µm. (G) Measurements of the area stained with F4/80 antibody in the crown-like structures of adipose tissues from the experimental mice in (A). WAT, epididymal white adipose tissue; SC, subcutaneous fat; NS, non-significant. aP<0.05, bP<0.01, cP<0.001.
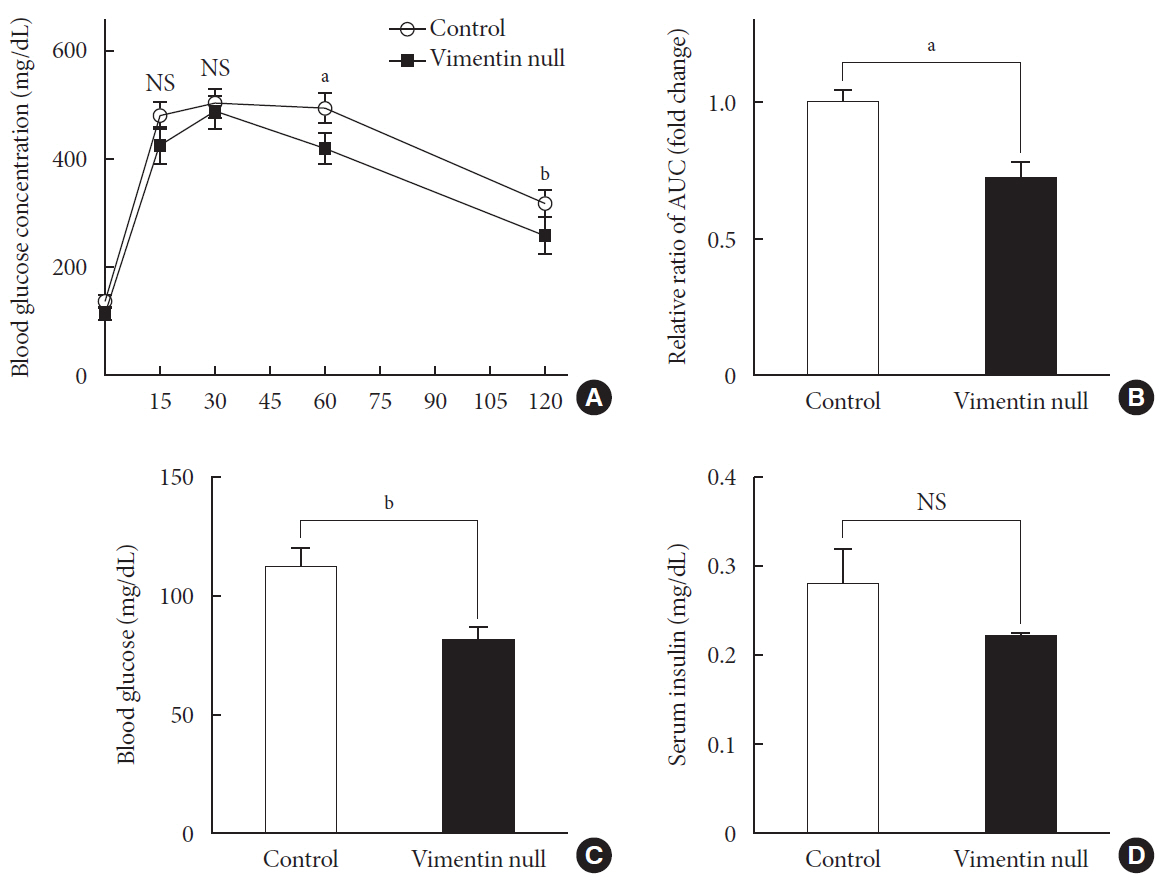
Fig. 2 Vim−/− mice exhibit enhanced glucose tolerance and lower fasting glucose. (A) Glucose tolerance after a 10-week high-fat diet (n=8 for control, n=8 for Vim−/− mice). (B) Area under the curve (AUC) of (A). (C) Blood glucose levels after 16 hours of fasting (n=10 for control, n=10 for Vim−/− mice). (D) Serum insulin concentrations after 16 hours of fasting (n=6 for control, n=6 for Vim−/− mice). The graph shows mean±standard error of the mean). NS, non-significant. aP<0.01, bP<0.001.
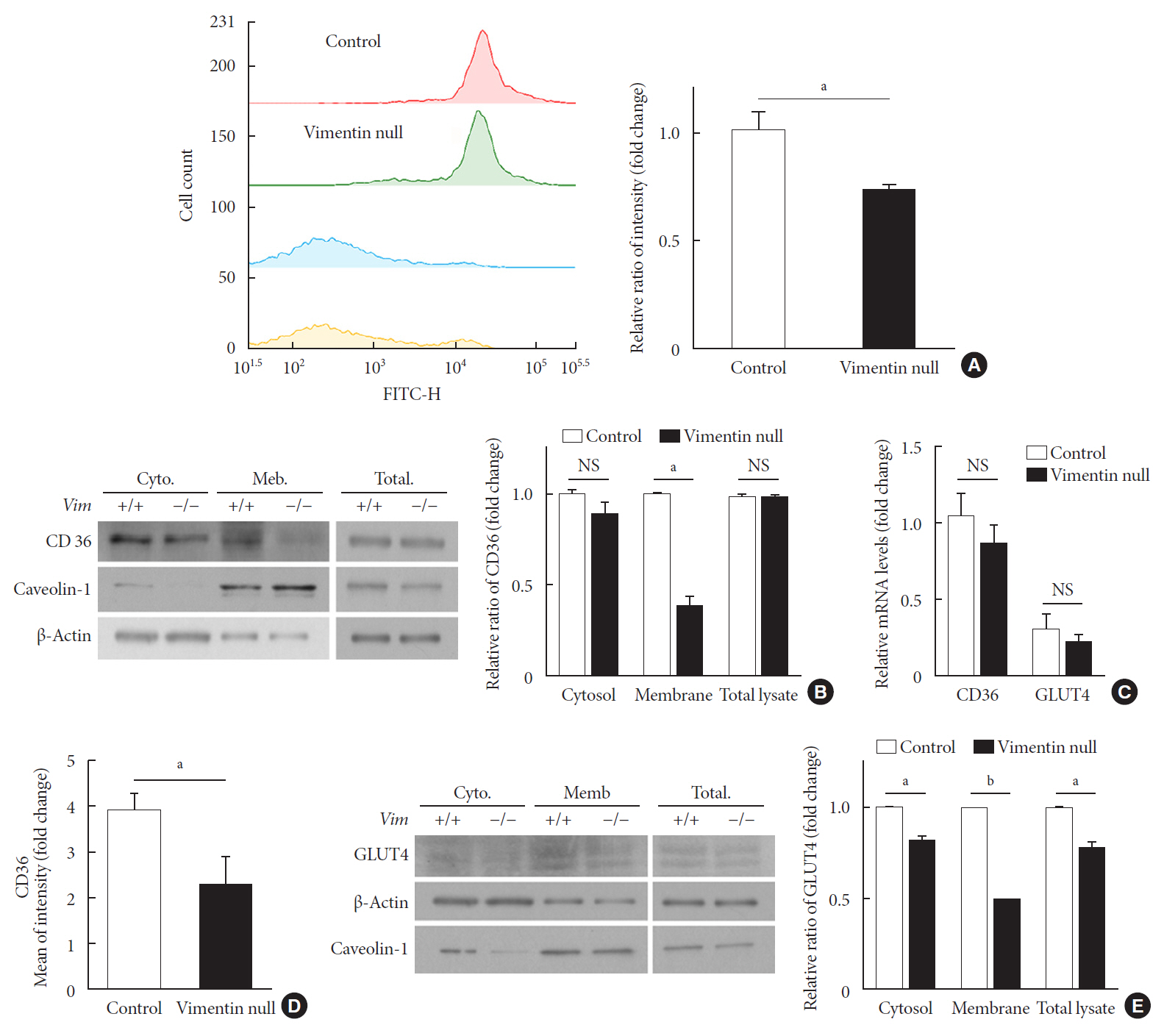
Fig. 3 Vimentin deficiency reduces CD36 and glucose transporter type 4 (GLUT4) trafficking to the plasma membrane in adipocytes. (A) Representative flow cytometry data of control and Vim−/− mouse adipocytes (left panel). Adipocytes were exposed to TF2-C12 fatty acid 40 µg/mL for 30 minutes. Fluorescence intensity of these cells was measured using flow cytometry (red: control+TF2-C12, green: Vim−/−+TF2-C12, light blue: untreated control adipocytes, yellow: untreated vimentin-null adipocytes). Geometric mean fluorescence intensity (right panel). (B) Western blot analysis for CD36 using the fractionated lysate from control and Vim−/− murine adipocytes. Cytosolic and membrane fractions were separated using a buffer-based protocol. Caveolin-1 was used as a marker for the plasma membrane fraction, and β-actin was used as a marker for the cytosolic fraction. (C) Quantitative real-time polymerase chain reaction to measure CD36 and GLUT4 expression in control and Vim−/− murine adipocytes. The values are normalized to the expression level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). (D) Plasma membrane-localized CD36 was measured by flow cytometry using allophycocyanin (APC)-conjugated CD36 antibody. Mean fluorescence intensities representing CD36 expression were compared between control and Vim−/− adipocytes. (E) Western blot analysis for GLUT4 using the fractionated lysates from control and Vim−/− murine adipocytes. Cytosolic and membrane fractions were separated using a buffer-based protocol. Caveolin-1 was used as a marker for the membrane fraction, and β-actin was used as a marker for the cytosolic fraction. Quantitation of band intensities (right graph). Cyto, cytosol fraction; Mem., membrane fraction; NS, non-significant. aP<0.01, bP<0.001.
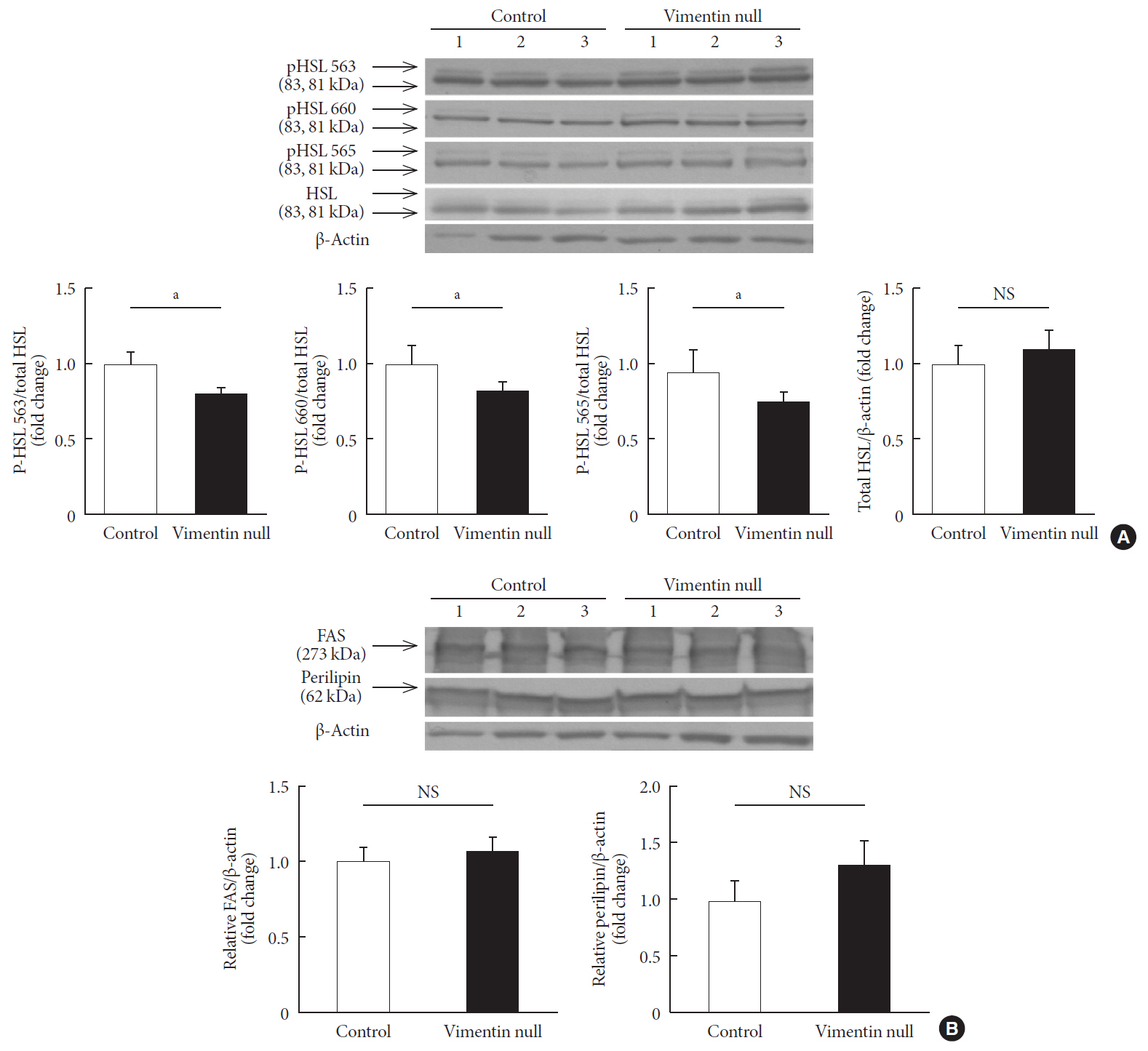
Fig. 4 Vimentin deficiency decreased lipolysis via hormone-sensitive lipase (HSL) activity. (A) Western blot analyses for HSL and phosphorylated HSL (p-HSL; Ser563, Ser660, Ser565) using homogenized epididymal fat lysates from the experimental mice in Fig. 1A (control, n=3; Vim−/−, n=3). Quantitation of the band intensities in Western blots (graphs below). (B) Western blot analyses for fatty acid synthase and perilipin using fat lysates from the experimental mice in Fig. 1A (control, n=3; Vim−/−, n=3). Quantitation of the band intensities in Western blots (graphs below). The graph shows mean±standard error of the mean. FAS, fatty acid synthase; NS, non-significant. aP<0.05.
Cited by 2 articles
-
The Role of Adipose Tissue Lipolysis in Diet-Induced Obesity: Focus on Vimentin
Eun Roh, Hye Jin Yoo
Diabetes Metab J. 2021;45(1):43-45. doi: 10.4093/dmj.2020.0293.Extracellular Vimentin Alters Energy Metabolism And Induces Adipocyte Hypertrophy
Ji-Hae Park, Soyeon Kwon, Young Mi Park
Diabetes Metab J. 2024;48(2):215-230. doi: 10.4093/dmj.2022.0332.
Reference
-
1. GBD 2015 Obesity Collaborators. Afshin A, Forouzanfar MH, Reitsma MB, Sur P, Estep K, et al. Health effects of overweight and obesity in 195 countries over 25 years. N Engl J Med. 2017; 377:13–27.
Article2. Ye J. Mechanisms of insulin resistance in obesity. Front Med. 2013; 7:14–24.
Article3. Guo M, Ehrlicher AJ, Mahammad S, Fabich H, Jensen MH, Moore JR, et al. The role of vimentin intermediate filaments in cortical and cytoplasmic mechanics. Biophys J. 2013; 105:1562–1568.
Article4. Biskou O, Casanova V, Hooper KM, Kemp S, Wright GP, Satsangi J, et al. The type III intermediate filament vimentin regulates organelle distribution and modulates autophagy. PLoS One. 2019; 14:e0209665.
Article5. Liu CY, Lin HH, Tang MJ, Wang YK. Vimentin contributes to epithelial-mesenchymal transition cancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget. 2015; 6:15966–15983.
Article6. Heid H, Rickelt S, Zimbelmann R, Winter S, Schumacher H, Dorflinger Y, et al. On the formation of lipid droplets in human adipocytes: the organization of the perilipin-vimentin cortex. PLoS One. 2014; 9:e90386.
Article7. Shen WJ, Patel S, Eriksson JE, Kraemer FB. Vimentin is a functional partner of hormone sensitive lipase and facilitates lipolysis. J Proteome Res. 2010; 9:1786–1794.
Article8. Hirata Y, Hosaka T, Iwata T, Le CT, Jambaldorj B, Teshigawara K, et al. Vimentin binds IRAP and is involved in GLUT4 vesicle trafficking. Biochem Biophys Res Commun. 2011; 405:96–101.
Article9. Wilhelmsson U, Stillemark-Billton P, Boren J, Pekny M. Vimentin is required for normal accumulation of body fat. Biol Chem. 2019; 400:1157–1162.
Article10. Winzell MS, Ahren B. The high-fat diet-fed mouse: a model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes. 2004; 53 Suppl 3:S215–S219.
Article11. Longo M, Zatterale F, Naderi J, Parrillo L, Formisano P, Raciti GA, et al. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int J Mol Sci. 2019; 20:E2358.
Article12. Park YM. CD36, a scavenger receptor implicated in atherosclerosis. Exp Mol Med. 2014; 46:e99.
Article13. Govers R. Molecular mechanisms of GLUT4 regulation in adipocytes. Diabetes Metab. 2014; 40:400–410.
Article14. Krintel C, Morgelin M, Logan DT, Holm C. Phosphorylation of hormone-sensitive lipase by protein kinase A in vitro promotes an increase in its hydrophobic surface area. FEBS J. 2009; 276:4752–4762.15. Colucci-Guyon E, Portier MM, Dunia I, Paulin D, Pournin S, Babinet C. Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell. 1994; 79:679–694.
Article16. Shen WJ, Zaidi SK, Patel S, Cortez Y, Ueno M, Azhar R, et al. Ablation of vimentin results in defective steroidogenesis. Endocrinology. 2012; 153:3249–3257.
Article17. Bjorndal B, Burri L, Staalesen V, Skorve J, Berge RK. Different adipose depots: their role in the development of metabolic syndrome and mitochondrial response to hypolipidemic agents. J Obes. 2011; 2011:490650.18. Pepino MY, Love-Gregory L, Klein S, Abumrad NA. The fatty acid translocase gene CD36 and lingual lipase influence oral sensitivity to fat in obese subjects. J Lipid Res. 2012; 53:561–566.
Article19. Febbraio M, Silverstein RL. CD36: implications in cardiovascular disease. Int J Biochem Cell Biol. 2007; 39:2012–2030.
Article20. Oquendo P, Hundt E, Lawler J, Seed B. CD36 directly mediates cytoadherence of plasmodium falciparum parasitized erythrocytes. Cell. 1989; 58:95–101.
Article21. Griffin E, Re A, Hamel N, Fu C, Bush H, McCaffrey T, et al. A link between diabetes and atherosclerosis: glucose regulates expression of CD36 at the level of translation. Nat Med. 2001; 7:840–846.
Article22. Rac ME, Safranow K, Poncyljusz W. Molecular basis of human CD36 gene mutations. Mol Med. 2007; 13:288–296.
Article23. Nagy L, Tontonoz P, Alvarez JG, Chen H, Evans RM. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARgamma. Cell. 1998; 93:229–240.24. Febbraio M, Abumrad NA, Hajjar DP, Sharma K, Cheng W, Pearce SF, et al. A null mutation in murine CD36 reveals an important role in fatty acid and lipoprotein metabolism. J Biol Chem. 1999; 274:19055–19062.
Article25. Abdoul-Azize S, Atek-Mebarki F, Bitam A, Sadou H, Koceir EA, Khan NA. Oro-gustatory perception of dietary lipids and calcium signaling in taste bud cells are altered in nutritionally obesity-prone Psammomys obesus. PLoS One. 2013; 8:e68532.
Article26. Hajri T, Han XX, Bonen A, Abumrad NA. Defective fatty acid uptake modulates insulin responsiveness and metabolic responses to diet in CD36-null mice. J Clin Invest. 2002; 109:1381–1389.
Article27. Furuhashi M, Ura N, Nakata T, Shimamoto K. Insulin sensitivity and lipid metabolism in human CD36 deficiency. Diabetes Care. 2003; 26:471–474.
Article28. Nath A, Li I, Roberts LR, Chan C. Elevated free fatty acid uptake via CD36 promotes epithelial-mesenchymal transition in hepatocellular carcinoma. Sci Rep. 2015; 5:14752.
Article29. Chang L, Chiang SH, Saltiel AR. Insulin signaling and the regulation of glucose transport. Mol Med. 2004; 10:65–71.
Article30. Asano T, Fujishiro M, Kushiyama A, Nakatsu Y, Yoneda M, Kamata H, et al. Role of phosphatidylinositol 3-kinase activation on insulin action and its alteration in diabetic conditions. Biol Pharm Bull. 2007; 30:1610–1616.
Article31. Huang S, Czech MP. The GLUT4 glucose transporter. Cell Metab. 2007; 5:237–252.
Article32. Carvalho E, Kotani K, Peroni OD, Kahn BB. Adipose-specific overexpression of GLUT4 reverses insulin resistance and diabetes in mice lacking GLUT4 selectively in muscle. Am J Physiol Endocrinol Metab. 2005; 289:E551–E561.
Article33. Atkinson BJ, Griesel BA, King CD, Josey MA, Olson AL. Moderate GLUT4 overexpression improves insulin sensitivity and fasting triglyceridemia in high-fat diet-fed transgenic mice. Diabetes. 2013; 62:2249–2258.
Article34. Roefs MM, Carlotti F, Jones K, Wills H, Hamilton A, Verschoor M, et al. Increased vimentin in human α- and β-cells in type 2 diabetes. J Endocrinol. 2017; 233:217–227.
Article35. Kumar N, Robidoux J, Daniel KW, Guzman G, Floering LM, Collins S. Requirement of vimentin filament assembly for beta3-adrenergic receptor activation of ERK MAP kinase and lipolysis. J Biol Chem. 2007; 282:9244–9250.36. Asensio C, Jimenez M, Kuhne F, Rohner-Jeanrenaud F, Muzzin P. The lack of beta-adrenoceptors results in enhanced insulin sensitivity in mice exhibiting increased adiposity and glucose intolerance. Diabetes. 2005; 54:3490–3495.37. Potokar M, Kreft M, Li L, Daniel Andersson J, Pangrsic T, Chowdhury HH, et al. Cytoskeleton and vesicle mobility in astrocytes. Traffic. 2007; 8:12–20.
Article38. Samovski D, Sun J, Pietka T, Gross RW, Eckel RH, Su X, et al. Regulation of AMPK activation by CD36 links fatty acid uptake to β-oxidation. Diabetes. 2015; 64:353–359.
Article39. Zhou D, Samovski D, Okunade AL, Stahl PD, Abumrad NA, Su X. CD36 level and trafficking are determinants of lipolysis in adipocytes. FASEB J. 2012; 26:4733–4742.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Blood Sugar Control and Low-Carbohydrate High-Fat Diet
- Anti-obesity and LDL-cholesterol lowering effects of silkworm hemolymph in C57BL/6N mice fed high fat diet
- Ameliorative effect of myricetin on insulin resistance in mice fed a high-fat, high-sucrose diet
- Sasa borealis leaves extract improves insulin resistance by modulating inflammatory cytokine secretion in high fat diet-induced obese C57/BL6J mice
- Effect of Exercise Training on Insulin Sensitivity and Intracellular Glucose Metabolism in Skeletal Muscle of High Fat-fed Rats
