Calcium-sensing receptor-mediated L-tryptophan-induced secretion of cholecystokinin and glucose-dependent insulinotropic peptide in swine duodenum
- Affiliations
-
- 1Laboratory of Gastrointestinal Microbiology, Nanjing Agricultural University, Nanjing 210095, China. suqinhang69@njau.edu.cn
- KMID: 2407617
- DOI: http://doi.org/10.4142/jvs.2018.19.2.179
Abstract
- This study aimed to elucidate the effect of tryptophan (Trp) on gut hormone secretion as well as the roles of the calcium-sensing receptor (CaSR) and its downstream signaling pathway in gut hormone secretion by assessing swine duodenal perfusion in vitro. Swine duodenum was perfused with Krebs-Henseleit buffer as a basal solution. Various concentrations (0, 10, and 20 mM) of Trp were applied to investigate its effect on gut hormone secretion. A CaSR antagonist was used to detect the involvement of CaSR and its signal molecules. The 20 mM Trp concentration promoted the secretion of cholecystokinin (CCK) and glucose-dependent insulinotropic peptide (GIP), elevated the mRNA level of CaSR, and upregulated the protein levels of CaSR, protein kinase C (PKC), and inositol trisphosphate receptor (IP3R). However, NPS 2143, an inhibitor of CaSR, attenuated the CCK and GIP release, reduced the mRNA level of CaSR, and decreased the protein levels of CaSR, PKC, and IP3R with 20 mM Trp perfusion. The results indicate that CCK and GIP secretion can be induced by Trp in swine duodenum in vitro, and the effect is mediated by CaSR and its downstream signal molecules PKC and IP3R.
MeSH Terms
Figure
-
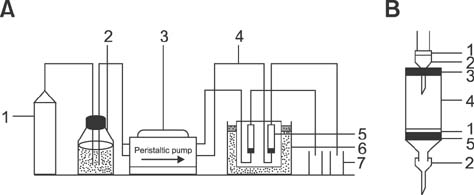
Fig. 1 Schematic of the perfusion system (A) and the perfusion chamber (B). In (A): 1, oxygen bomb (mixture of 95% O2 and 5% CO2); 2, media reservoir; 3, peristaltic pump; 4, silicone tube; 5, perfusion chamber; 6, water bath (37℃); 7, collector. In (B): 1, nylon filter platform; 2, needle; 3, rubber stopper; 4, syringe; 5, rubber plunger.

Fig. 2 The mRNA expression of CaSR (A) and the secretion of gut hormones (B and C) regulated by different concentrations of tryptophan (Trp). The duodenal tissues were perfused with 0 (control), 10, and 20 mM Trp for 120 min. Tissues were collected at the end of perfusion. CaSR (A) mRNA expression was analyzed by quantitative polymerase chain reaction (qPCR) and glyceraldehyde phosphate dehydrogenase was used as the internal control. The concentrations of cholecystokinin (CCK, B) and glucose-dependent insulinotropic peptide (GIP, C) in the perfusate samples were detected every 20 min by using enzyme-linked immunosorbent assay kits. Values are expressed as mean ± SEM (n = 4). A statistical difference was determined by one-way ANOVA followed by Tukey's multiple comparison tests. Bars with the different letters show significant differences compared with the control group (A) (p < 0.05); * and δ (B and C); 20 mM Trp and 10 mM Trp treatments are significantly different from the control, respectively (p < 0.05). The influence of Ca2+ on the ability of Trp to induce the expression of CaSR (D) and the secretion of gut hormones (E and F). Tissue samples of duodenum were perfused with Krebs-Henseleit buffer containing 0 mM Trp without [Ca2+]e (control) and 20 mM Trp with or without 10 mM [Ca2+]e for 120 min. After perfusion, the expression of CaSR (D) was determined by qPCR. Perfusate samples were collected every 20 min to measure concentrations of CCK (E) and GIP (F). Values are expressed as mean ± SEM (n = 4). Statistical differences were determined by one-way ANOVA followed by Tukey's multiple comparison tests. Bars with the different letters showed the significant differences compared with the control group (D) (p < 0.05); γ (E and F); the combination of 20 mM Trp and 10 mM Ca2+ was significantly different from the control (p < 0.05). Effects of CaSR antagonist NPS 2143 on the expression of Trp-induced CaSR (G) and the secretion of gut hormones (H and I). Duodenal tissues were treated with 0 (control), 20, or 20 mM Trp and NPS 2143 (25 µM). At the end of perfusion (120 min), the duodenum tissues were collected to determine the mRNA expression of CaSR (G) by qPCR. The perfusate samples were obtained every 20 min to evaluate the concentration of CCK (H) and GIP (I). Values are expressed as mean ± SEM (n = 6). Statistical differences were determined by one-way ANOVA followed by Tukey's multiple comparison tests. Bars with the different letters showed the significant differences compared with the control group (G) (p < 0.05); χ (H and I); 20 mM Trp and the combination of 20 mM Trp and NPS 2143 treatments are significantly different from the control, respectively (p < 0.05).

Fig. 3 Protein levels of calcium-sensing receptor (CaSR) and its signal molecules regulated by tryptophan (Trp). (A) The protein images of CaSR, protein kinase C (PKC), and inositol trisphosphate receptor (IP3R) at 0 and 20 mM Trp treatments. The optical densities of CaSR, PKC, and IP3R are shown in (B–D), respectively. Values are shown as mean ± SEM. **p < 0.01.

Fig. 4 The protein levels of calcium-sensing receptor (CaSR) and its signal molecules regulated by CaSR antagonist NPS 2143. (A) The protein images of CaSR, protein kinase C (PKC), and inositol trisphosphate receptor (IP3R) in the presence of 20 mM tryptophan and NPS 2143. The optical densities of CaSR, PKC, and IP3R are shown in (B–D), respectively. Values are shown as mean ± SEM. **p < 0.01.
Reference
-
1. Badger TM. Perifusion of anterior pituitary cells: release of gonadotropins and somatotropins. Methods Enzymol. 1986; 124:79–90.2. Bronk JR, Hastewell JG. The transport and metabolism of naturally occurring pyrimidine nucleosides by isolated rat jejunum. J Physiol. 1988; 395:349–361.
Article3. Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, Vandesompele J, Wittwer CT. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009; 55:611–622.
Article4. Conigrave AD, Brown EM. Taste receptors in the gastrointestinal tract. II. L-amino acid sensing by calcium-sensing receptors: implications for GI physiology. Am J Physiol Gastrointest Liver Physiol. 2006; 291:G753–G761.5. Conigrave AD, Franks AH, Brown EM, Quinn SJ. L-amino acid sensing by the calcium-sensing receptor: a general mechanism for coupling protein and calcium metabolism? Eur J Clin Nutr. 2002; 56:1072–1080.
Article6. Cuber JC, Bernard C, Levenez F, Chayvialle JA. [Lipids, proteins and carbohydrates stimulate the secretion of intestinal cholecystokinin in the pig]. Reprod Nutr Dev. 1990; 30:267–275. French.7. Hansen JR, Conn PM. Measurement of pulsatile hormone release from perifused pituitary cells immobilized on microcarriers. In : Conn PM, editor. Methods in Neurosciences. Vol. 2. Cell Culture. San Diego: Academic Press;1990. p. 181–194.8. Hebert SC, Cheng S, Geibel J. Functions and roles of the extracellular Ca2+-sensing receptor in the gastrointestinal tract. Cell Calcium. 2004; 35:239–247.
Article9. Hira T, Nakajima S, Eto Y, Hara H. Calcium-sensing receptor mediates phenylalanine-induced cholecystokinin secretion in enteroendocrine STC-1 cells. FEBS J. 2008; 275:4620–4626.
Article10. Hofer AM, Brown EM. Extracellular calcium sensing and signalling. Nat Rev Mol Cell Biol. 2003; 4:530–538.
Article11. Hou G, Xue L, Lu Z, Fan T, Tian F, Xue Y. An activated mTOR/p70S6K signaling pathway in esophageal squamous cell carcinoma cell lines and inhibition of the pathway by rapamycin and siRNA against mTOR. Cancer Lett. 2007; 253:236–248.
Article12. Karhunen LJ, Juvonen KR, Huotari A, Purhonen AK, Herzig KH. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul Pept. 2008; 149:70–78.
Article13. Liddle RA, Green GM, Conrad CK, Williams JA. Proteins but not amino acids, carbohydrates, or fats stimulate cholecystokinin secretion in the rat. Am J Physiol. 1986; 251:G243–G248.
Article14. Liou AP, Sei Y, Zhao X, Feng J, Lu X, Thomas C, Pechhold S, Raybould HE, Wank SA. The extracellular calcium-sensing receptor is required for cholecystokinin secretion in response to L-phenylalanine in acutely isolated intestinal I cells. Am J Physiol Gastrointest Liver Physiol. 2011; 300:G538–G546.
Article15. Mace OJ, Schindler M, Patel S. The regulation of K- and L-cell activity by GLUT2 and the calcium-sensing receptor CasR in rat small intestine. J Physiol. 2012; 590:2917–2936.
Article16. Macleod RJ. CaSR function in the intestine: hormone secretion, electrolyte absorption and secretion, paracrine non-canonical Wnt signaling and colonic crypt cell proliferation. Best Pract Res Clin Endocrinol Metab. 2013; 27:385–402.
Article17. Miazza B, Palma R, Lachance JR, Chayvialle JA, Jonard PP, Modigliani R. Jejunal secretory effect of intraduodenal food in humans. A comparison of mixed nutrients, proteins, lipids, and carbohydrates. Gastroenterology. 1985; 88:1215–1222.
Article18. Moran-Ramos S, Tovar AR, Torres N. Diet: friend or foe of enteroendocrine cells--how it interacts with enteroendocrine cells. Adv Nutr. 2012; 3:8–20.
Article19. Murphy KG, Bloom SR. Gut hormones and the regulation of energy homeostasis. Nature. 2006; 444:854–859.
Article20. Neary MT, Batterham RL. Gut hormones: implications for the treatment of obesity. Pharmacol Ther. 2009; 124:44–56.
Article21. Nemeth EF, Steffey ME, Hammerland LG, Hung BC, Van Wagenen BC, DelMar EG, Balandrin MF. Calcimimetics with potent and selective activity on the parathyroid calcium receptor. Proc Natl Acad Sci U S A. 1998; 95:4040–4045.
Article22. Rasoamanana R, Darcel N, Fromentin G, Tomé D. Nutrient sensing and signalling by the gut. Proc Nutr Soc. 2012; 71:446–455.
Article23. Steinert RE, Beglinger C. Nutrient sensing in the gut: interactions between chemosensory cells, visceral afferents and the secretion of satiation peptides. Physiol Behav. 2011; 105:62–70.
Article24. Steinert RE, Luscombe-Marsh ND, Little TJ, Standfield S, Otto B, Horowitz M, Feinle-Bisset C. Effects of intraduodenal infusion of L-tryptophan on ad libitum eating, antropyloroduodenal motility, glycemia, insulinemia, and gut peptide secretion in healthy men. J Clin Endocrinol Metab. 2014; 99:3275–3284.
Article25. Tan YF, Chen WH, Zou SX. The effect of β Casomorphin on piglets' stomach antrum under superfusion. J Nanjing Agric Univ. 2000; 23:72–75.26. Wang Y, Chandra R, Samsa LA, Gooch B, Fee BE, Cook JM, Vigna SR, Grant AO, Liddle RA. Amino acids stimulate cholecystokinin release through the Ca2+-sensing receptor. Am J Physiol Gastrointest Liver Physiol. 2011; 300:G528–G537.
Article27. Zhou HR, Pestka JJ. Deoxynivalenol (vomitoxin)-induced cholecystokinin and glucagon-like peptide-1 release in the STC-1 enteroendocrine cell model is mediated by calcium-sensing receptor and transient receptor potential ankyrin-1 channel. Toxicol Sci. 2015; 145:407–417.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- The Role of the Sweet Taste Receptor in Enteroendocrine Cells and Pancreatic beta-Cells
- The Incretins and Pancreatic beta-Cells: Use of Glucagon-Like Peptide-1 and Glucose-Dependent Insulinotropic Polypeptide to Cure Type 2 Diabetes Mellitus
- The Cardiovascular Effect of Tirzepatide: A Glucagon-Like Peptide-1 and Glucose-Dependent Insulinotropic Polypeptide Dual Agonist
- Calcium Sensing Receptor
- Incretin Hormones: Pathophysiological Risk Factors and Potential Targets for Type 2 Diabetes
