Early cerebellar granule cell migration in the mouse embryonic development
- Affiliations
-
- 1Department of Cell Biology and Human Anatomy, School of Medicine, University of California, Davis, Sacramento, California 95817, USA.
- 2Department of Anatomy, College of Medicine, Konyang University, Daejeon, Korea. ygjeong@konyang.ac.kr
- 3Department of Companion Animal and Animal Resources Science, Joongbu University, Geumsan-gun, Korea.
- KMID: 2168901
- DOI: http://doi.org/10.5115/acb.2010.43.1.86
Abstract
- Pax6, a paired homeobox DNA binding protein, has been found to be expressed in the cerebellum in both granule cells and their precursors in the external granular layer (EGL). In this study we have traced Pax6 expression through embryonic development in mice by using a polyclonal antibody against Pax6 and used it to study the cellular dispersal pattern of the EGL. During dispersal the EGL was thicker and Pax6 expression was more intense on the rostral side of the lateral corners of the cerebellum. Pax6 immunoreactive cells were found to be migrating from the EGL during the early stage of EGL dispersal, which suggested the early inward migration of granule cells. Double staining with various markers confirmed that the early-migrating cells are not Purkinje cells, interneurons or glia. Although the Pax6 immunoreactive cells within the cerebellum were not apparently proliferating, NeuN, a marker for postmitotic granule cells, was not expressed in these cells until E16. Furthermore, granule cells were observed migrating inwards from the EGL both during and after EGL dispersal. These early migrating granule cells populated the whole cerebellum. These findings offer novel views on specific stages of granule cell dispersal and migration.
Keyword
MeSH Terms
Figure
-
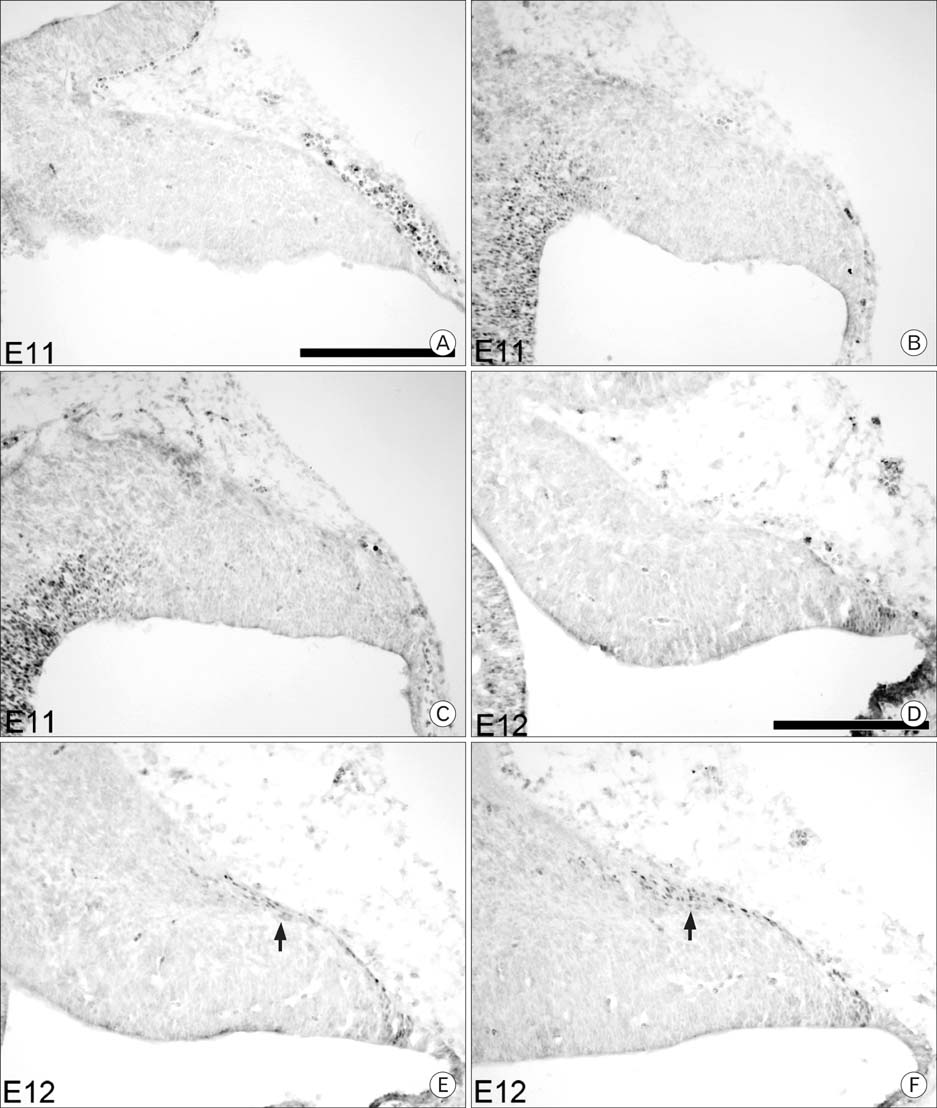
Fig. 1 Pax6 expression at embryonic day E11 and E12 in the cerebellum. The top row shows medial sections, the middle row shows paramedial sections and the bottom row shows lateral sections that correspond to mediolateral locations. Immunocytochemistry with an antibody against Pax6 was performed on sagittal sections. No Pax6 expression is observed at E11 (A~C). At E12 (D~F), cells immunoreactive for Pax6 are first seen in a single layer on the dorsal surface, as well as a small group of cells at the rostral end of the layer (arrow). Scale bar is 250 µm.
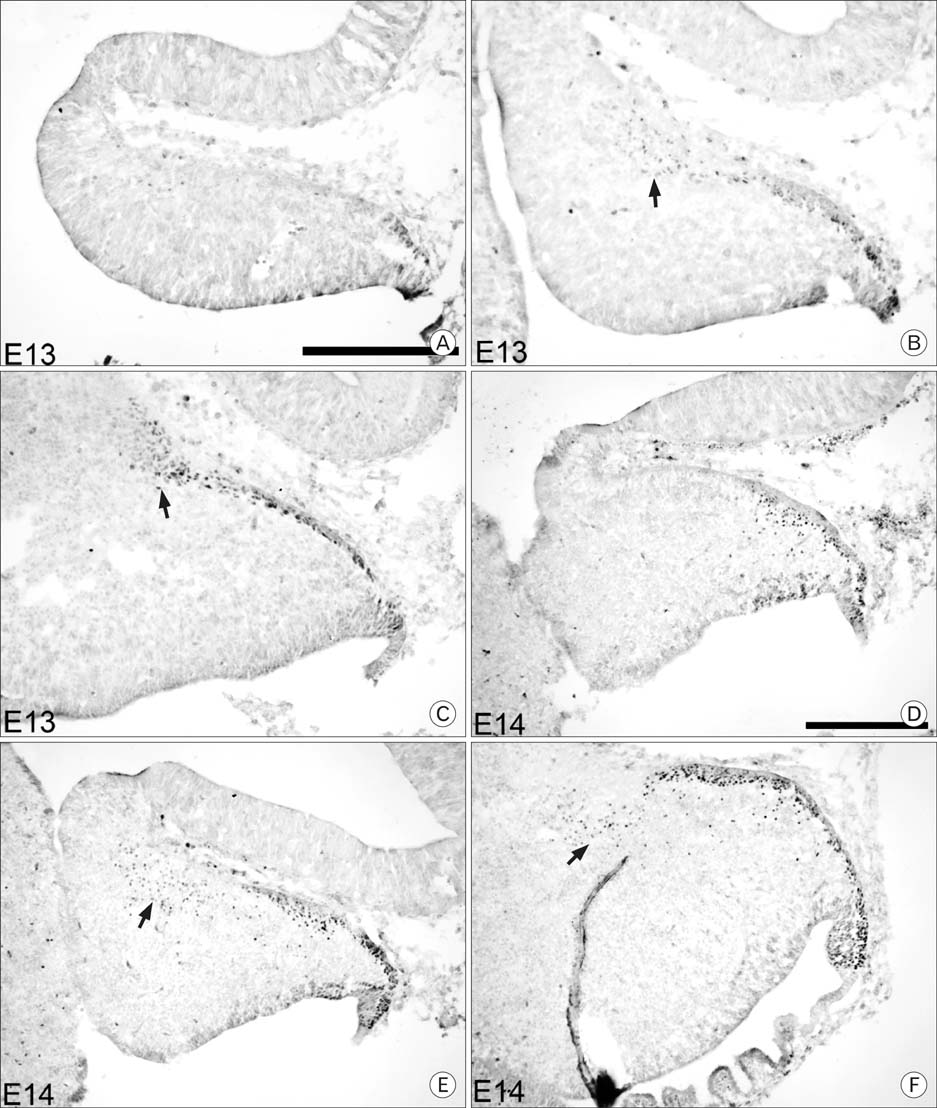
Fig. 2 Pax6 expression in E13 and E14 cerebella. At E13 (A~C), the dorsal layer has increased, and the rostral group has increased in size and number. At E14 (D~F), the EGL is first observed in the medial sections, and inward migration has begun. Ed-The authors should state what each row of images is referring to, even if it is the same as in Fig. 1. Also, the authors should state what the arrows are pointing to in panels (B, C, E, F) Scale bar is 250 µm.

Fig. 3 Pax6 expression in E15 and E16 cerebella. At E15 (A~C), the rostral group of cells, as well as all other Pax6 immunoreactive cells within the cerebellum, has reached the ventral side of the cerebellum in the paramedial sections, and a discontinuity in EGL thickness is observed in the dorsocaudal corner of the cerebellum in the lateral sections (arrowheads). At E16 (D~F), this discontinuity is more obvious, and the rostral group of cells no longer expresses Pax6 and may have migrated caudally into the center of the cerebellum (asterisk). Ed-Similar to notes for Fig. 2, the authors should state what each row of images is referring to. Also, the authors should state what the arrows are pointing to. Scale bar is 250 µm.

Fig. 4 Pax6 expression in E17 and E18 cerebella. At E17 (A~C) and E18 (D~F), a rostral group of Pax6 immunoreactive cells is only seen in lateral sections. From E15 to E18, the discontinuity remains in the same rostrocaudal location. Arrows point to the rostral group of Pax6 immunoreactive cells. Arrowheads point to the rostral and caudal end of the discontinuity in the EGL of lobule VIb and VII. Asterisks indicate the presumed location of the rostral group of cells that were immunoreactive for Pax6 before E16. Panels A and D show medial sections, B and E show paramedial sections, and C and F show lateral sections. Scale bar is 250 µm.
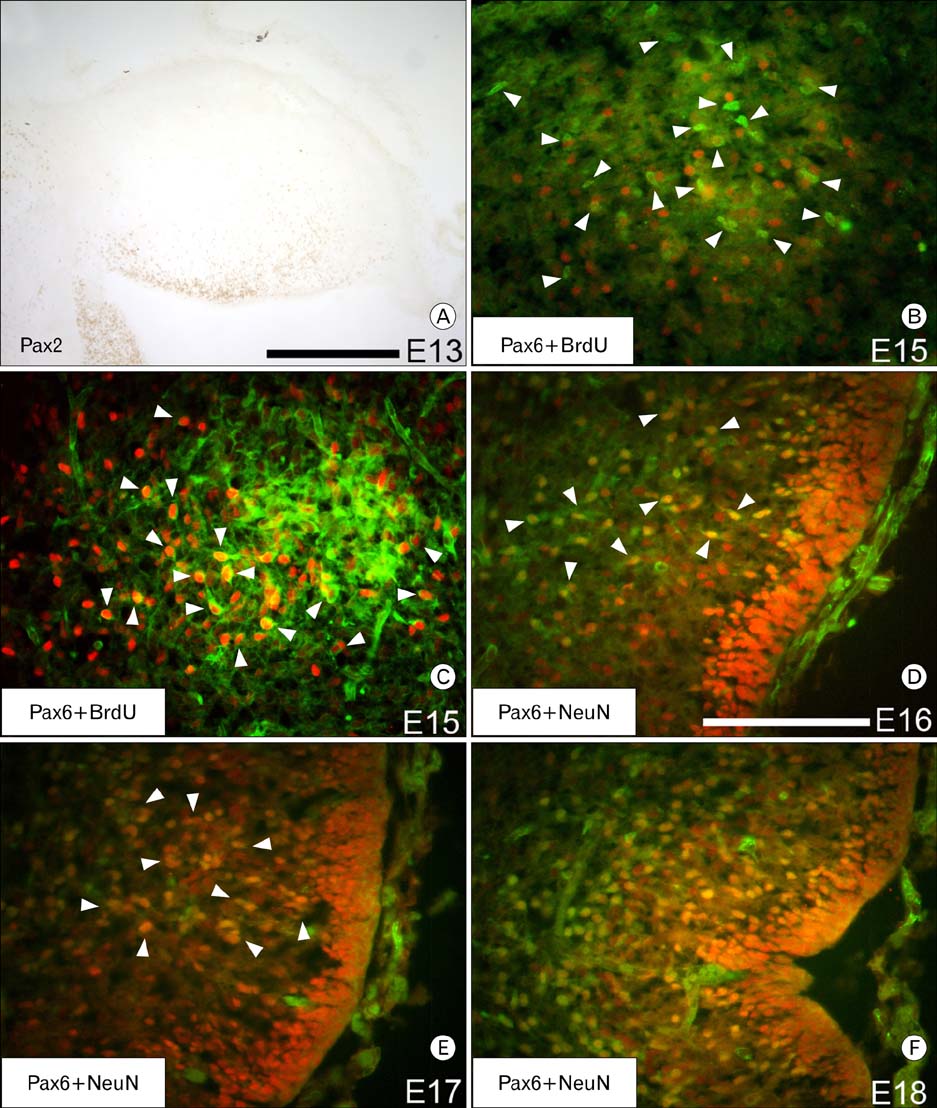
Fig. 5 Identification of the Pax6 immunoreactive cells within the cerebellum. Immunocytochemistry with an antibody against Pax2 shows a different migration pattern from Pax6 (A; rostral is left, dorsal is up). Double immunofluorescence labeling for BrdU (green) and Pax6 (red) of an E15 cerebellum that was injected with BrdU 10 minutes before sacrifice, shows all Pax6 immunoreactive cells within the cerebellum are postmitotic (B). Arrowheads in (B) point to cells that are immunoreactive for BrdU only. Double labeling with BrdU (green) and Pax6 (red) of an E15 cerebellum that was injected with BrdU at E10 shows that some cells of the rostral group of Pax6 immunoreactive cells are DCN cells (C). Arrowheads in (C) point to cells immunoreactive for both Pax6 and BrdU. Double immunofluorescence labeling for NeuN (green) and Pax6 (red) at E16 (D), E17 (E) and E18 (F) suggests that the Pax6 immunoreactive cells within the cerebellum are granule cells. Arrowheads in (D) and (E) point to cells immunoreactive for both Pax6 and NeuN. At E16, most cells are only immunoreactive for Pax6 (D). However, by E18 (F), all Pax6 immunoreactive cells within the posterior lobe of the cerebellum are also immunoreactive for NeuN. Scale bar in (A) is 250 µm; (D) is 125 µm. The scale for panel D also applies to (B~F).
Cited by 1 articles
-
A rare additional lobe of cerebellum, projecting from its superior surface
Satheesha Badagabettu Nayak, Suhani Sumalatha, Surekha Devadasa Shetty
Anat Cell Biol. 2022;55(3):376-379. doi: 10.5115/acb.22.048.
Reference
-
1. Altman J. Postnatal development of the cerebellar cortex in rat.I. The external germinal layer and the transitional molecular layer. J Comp Neurol. 1972. 145:353–397.2. Altman J, Bayer SA. Embryonic development of the rat cerebellum. II. Translocation and regional distribution of the deep neurons. J Comp Neurol. 1985. 231:27–41.3. Altman J, Bayer SA. Development of the cerebellar system: in relation to its evolution, structure, and functions. 1997. CRC Press;Chapter 9 and 10.4. Beierbach E. Lobulation in Mouse Cerebellum. 2001. University of Calgary, Canada: Masters thesis.5. Beimesche S, Neubauer A, Herzig S, et al. Tissue-specific transcriptional activity of a pancreatic islet cell-specific enhancer sequence/Pax6-binding site determined in normal adult tissues in vivo using transgenic mice. Mol Endocrinol. 1999. 13:718–728.6. Engelkamp D, Rashbass P, Seawright A, van Heyningen V. Role of Pax6 in development of the cerebellar system. Development. 1999. 126:3585–3596.7. Hanson I, Van Heyningen V. Pax6: more than meets the eye. Trends Genet. 1995. 11:268–272.8. Hanson IM, Seawright A, Hardman K, et al. Pax6 mutations in aniridia. Hum Mol Genet. 1993. 2:915–920.9. Hill RE, Favor J, Hogan BL, et al. Mouse small eye results from mutations in a paired-like homeobox-containing gene. Nature. 1991. 354:522–525.10. Jensen P, Smeyne R, Goldowitz D. Analysis of cerebellar development in math1 null embryos and chimeras. J Neurosci. 2004. 24:2202–2211.11. Karam SD, Kim YS, Bothwell M. Granule cells migrate within raphes in the developing cerebellum: an evolutionarily conserved morphogenic event. J Comp Neurol. 2001. 440:127–135.12. Kawano H, Fukuda T, Kubo K, et al. Pax-6 is required for thalamocortical pathway formation in fetal rats. J Comp Neurol. 1999. 408:147–160.13. Luckner R, Obst-Pernberg K, Hirano S, Suzuki ST, Redies C. Granule cell raphes in the developing mouse cerebellum. Cell Tissue Res. 2001. 303:159–172.14. Mastick GS, Davis NM, Andrew GL, Easter SS Jr. Pax6 functions in boundary formation and axon guidance in the embryonic mouse forebrain. Development. 1997. 124:1985–1997.15. Maricich SM, Herrup K. Pax-2 expression defines a subset of GABAergic interneurons and their precursors in the developing murine cerebellum. J Neurobiol. 1999. 41:281–294.16. Miller MW, Nowakowski RS. Use of bromodeoxyuridine-immunohistochemistry to examine the proliferation, migration and time of origin of cells in the central nervous system. Brain Res. 1988. 457:44–52.17. Weyer A, Schilling K. Developmental and cell type-specific expression of the neuronal marker NeuN in the murine cerebellum. J Neurosci Res. 2003. 73:400–409.18. Yamasaki T, Kawaji K, Ono K, et al. Pax6 regulates granule cell polarization during parallel fiber formation in the developing cerebellum. Development. 2001. 128:3133–3144.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Maternal effect genes: Findings and effects on mouse embryo development
- The Effects of Narcotics on the Mouse Two-Cell Embryo Development
- Effect of Cumulus Cell Coculture on Early Mouse Embryonal Development in vitro
- Effectsof Protein and Amino Acid(Glutamate) on the Development of Late 2-Cell Mouse Embryo in vitro
- Effect of Human Oviduct Epithelial Cells and Vero Cell on Early Mouse Embryonal Development In Vitro
