Stem Cells in Respiratory Diseases
- Affiliations
-
- 1Department of Internal Medicine, Soonchunhyang University Hospital, Bucheon, Korea. jas877@schbc.ac.kr
- KMID: 1910090
- DOI: http://doi.org/10.4046/trd.2007.63.2.121
Abstract
- No abstract available.
MeSH Terms
Figure
-
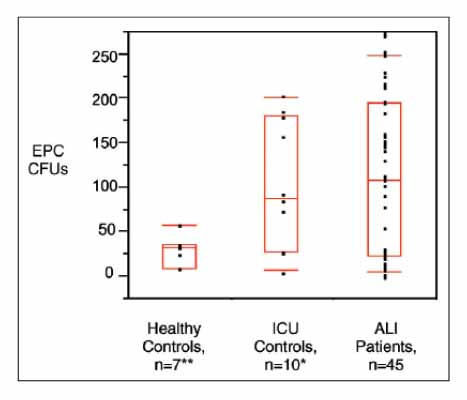
Figure 1 Numbers of EPC colony-forming units (CFUs) from healthy control patients, ICU control patients, and patients with ALI. Both ICU control patients and patients with ALI have significantly more EPC CFUs than do healthy control patients. *p=0.95 versus patients with ALI, **p<0.05 versus patients with ALI. Boxes represent the median, 25th,and 75th percentiles. The 10th and 90th percentiles are signified by floating red bars.
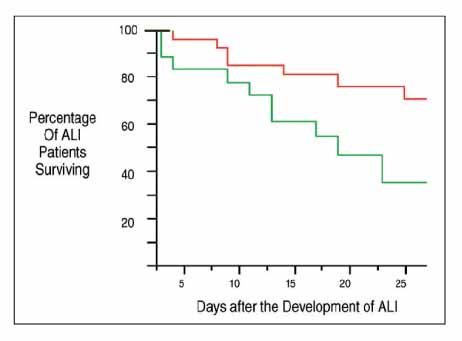
Figure 2 Kaplan-Meier curve of patients with ALI stratified by EPC CFU count of greater than or equal to 35 (red line) or less than 35 (green line). Patients with ALI with an EPC CFU count of greater than 35 had a survival benefit, with approximately 70% of these patients alive at 28 days, compared with 35% in those patients with EPC CFUs less than 35. p<0.03 for comparison between the two groups.
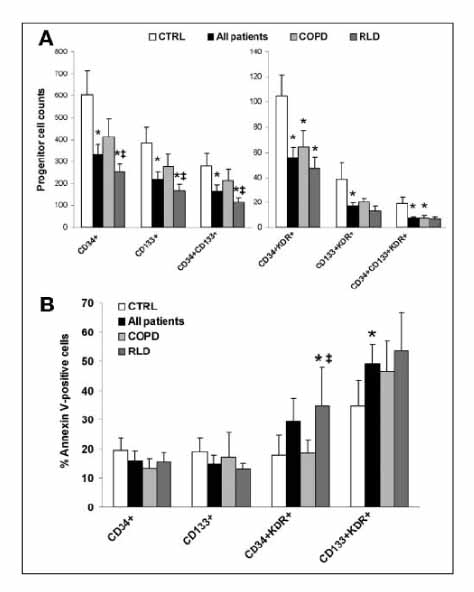
Figure 3 Reduction of circulating progenitor cells in patients with severe lung disease. (A): Comparisons of the levels of all progenitor cell subtypes in control subjects, in all patients with hypoxemia due to pulmonary disease, in patients with RLD, and in patients with COPD. Analysis of variance (ANOVA) p<.05 for all cell types. (B): Percentages of progenitor cells positive for Annexin V binding in the same four groups. ANOVA p<.05 for CD34 KDR and CD133 KDR cells. *p<.05 for t test comparing patients with controls. ‡p<.05 for t test comparing COPD with RLD patients. Abbreviations: COPD,chronic obstructive pulmonary disease; CTRL, control subjects; RLD, restrictive lung disease.
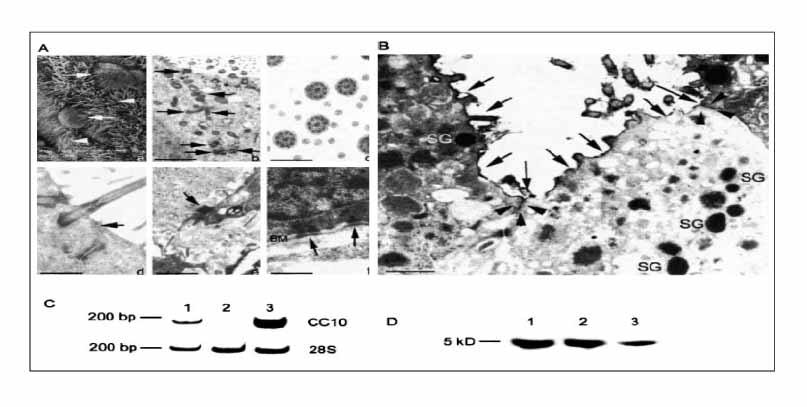
Figure 4 SEM of generated airway epithelium (Aa) showing ciliated cells (white arrowheads) and cells exhibiting a nonciliated bulging apical membrane, characteristic of Clara cells (white arrows). Ultrastructural features of in vitro ES-derived bioengineered airway epithelium observed by TEM (Ab, Ac, Ad, Ae, and Af). This epithelium exhibits active ciliogenesis demonstrated by centriole migration (black arrows) toward the apical plasma membrane (b) leading to the formation of mature cilia composed of nine tubule-pairs at the periphery and a tubule-doublet in the center of the cilia (c). The bioengineered epithelium exhibits tight junctions (black arrow) at the apical part of lateral plasma membranes (d), desmosomes (black arrow) at the lateral plasma membranes (e) and hemidesmosomes (black arrows) connecting the basal plasma membranes to the underlying basement membrane (BM) (f). Bars 10 µm (a); 150 nm (b), 30 nm (c), 75 nm (d), 295 nm (e), and 50 nm (f). (B) Functionality of the tight junctions analyzed using the lanthanum nitrate diffusion technique and visualized by TEM. Ultrathin sections show that lanthanum nitrate remains located at the apical surface of the ES-derived bioengineered airway epithelium (black arrows) and does not penetrate through the tight junctions (black arrowheads) between electron-dense secretory granule (SG)-containing Clara cells. Bar 50 nm. (C) Electrophoresis of RT-PCR products of CC10 mRNA extracted from control mouse trachea (lane 1), undifferentiated ES cells (lane 2) and ES-derived airway epithelium (lane 3) shows the expected 198-bp transcript in bioengineered airway epithelium. 28S housekeeping gene generates the expected 212-bp transcript. (D) Western blot analysis of the CC10 protein in the secretions covering the air-liquid interface cultures reveal a 5-kD protein corresponding to the CC10 protein (lane 3) also observed in native mouse lung (lane 1) and tracheal (lane 2) tissues
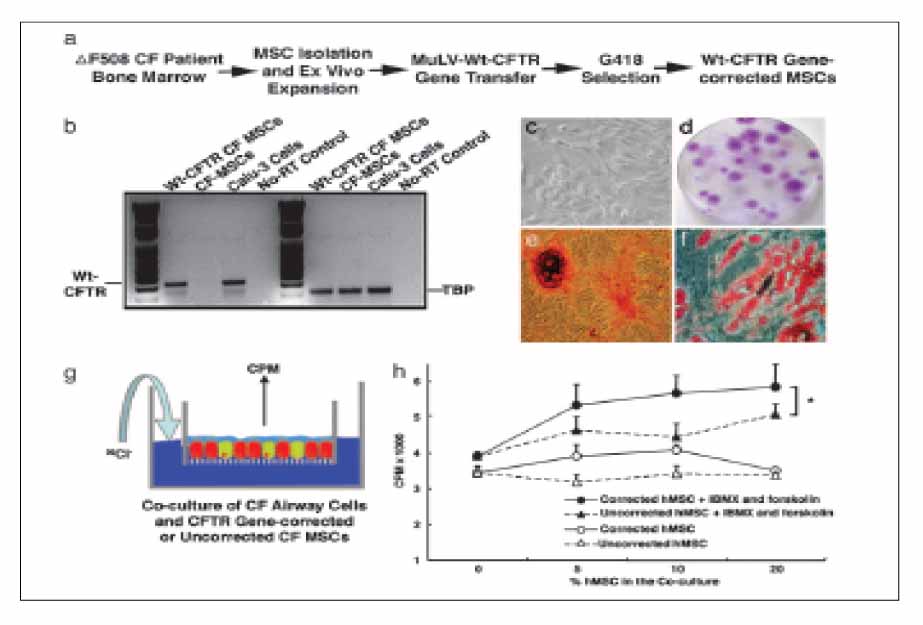
Figure 5 CFTR-corrected CF-patient MSCs retained their multipotency and responded to cAMP stimulation by secreting chloride to the apical side. (a) Schematic for CF-patient MSC isolation, expansion, gene correction, and positive drug selection. (b) RT-PCR to verify the successful CFTR gene transfer. RT-PCR was performed to amplify WT CFTR transcripts but not ΔF508 mutant transcripts. The gene-corrected CF MSCs and positive control Calu-3 cells have WT CFTR transcription, whereas non-gene-corrected CF MSCs and the no-RT control show negative amplification. In the TBP RT-PCR control, all of the samples except the no-RT control show positive PCR products. (c) Phase-contrast microscopic view of the CFTR gene-corrected CF-patient MSCs. (d) Photomicrograph of a representative stem cell colony plate. Purple-stained foci are the MSC colonies. (e) Osteogenesis of the CFTR gene-corrected CF-patient MSCs. After differentiation in an osteogenic medium, cells had mineral deposits visualized in red by Alizarin red staining. (f) Adipogenesis of the CFTR gene-corrected CF-patient MSCs. After differentiation in an adipogenic medium, cells had lipid droplet accumulation stained in red with oil red O. (g and h) CFTR gene-corrected MSCs from CF patients contributed to the apical cAMP-stimulated Cl- secretion. CFTR gene-corrected CF-patient MSCs or non-gene-corrected CF-patient MSCs were mixed with ΔF508 CF AECs at different ratios as indicated. After 1 month in culture at the air-liquid interface, chloride efflux assays were performed as described in Materials and Methods. A two-way ANOVA test revealed that cocultures with the CFTR gene-corrected CF-patient MSCs (F) had a greater chloride secretion in response to the IBMX and forskolin stimulation than the cocultures with non-gene-corrected CF-patient MSCs (OE) (n 4; P≤0.05).
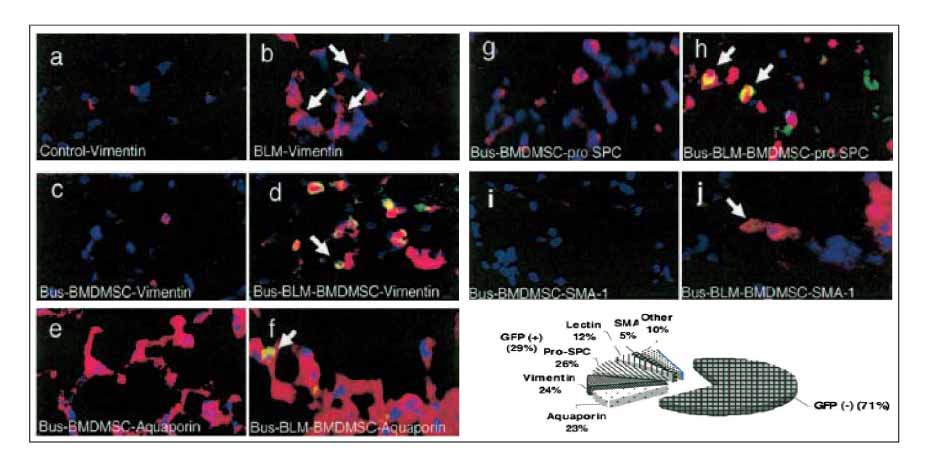
Figure 6 Donor BMDMSC localizing to injured lung assume lung cell phenotypes. Sections were analyzed in double-stained IIFA with anti-GFP (green) and antibodies to specific cell type markers (red); co-localization in each case appears yellow (arrows point to double positive cells). (a-d) Anti-vimentin (fibroblast). (a) Normal control; (b) 14 d after bleomycin; (c) 14 d after busulfan followed by BMDMSC (no lung injury); (d) 14 d after bleomycin followed by BMDMSC in a busulfan myelosuppressed animal. (e, f) Anti-aquaporin (type I alveolar epithelium). (e) Fourteen days after busulfan followed by BMDMSC (no lung injury); (f) 14 d after bleomycin followed by BMDMSC in busulfan myelosuppressed animal. (g, h) Anti-pro-surfactant protein C (type II alveolar epithelium). (g) Fourteen days after busulfan followed by BMDMSC (no lung injury); (h) 14 d after bleomycin followed by BMDMSC in busulfan myelosuppressed animal. (i, j) Anti-smooth muscle actin (SMA-1, myofibroblasts). (i) Fourteen days after busulfan followed by BMDMSC (no lung injury); (j) 14 d after bleomycin followed by BMDMSC in busulfan myelosuppressed animal. (k) Percentage of GFP-positive cells that express lung cell phenotype markers in myelosupressed mice treated with bleomycin and infused with BMDMSC. All microphotographs were taken at 40 magnification.
Reference
-
1. Sanders RC Jr, Slayton WB, Cogle CR, Fisher RC, Scott EW. Stem cell research. Paediatr Respir Rev. 2006. 7:135–140.2. Burnham EL, Taylor WR, Quyyumi AA, Rojas M, Brigham KL, Moss M. Increased circulating endothelial progenitor cells are associated with survival in acute lung injury. Am J Respir Crit Care Med. 2005. 172:854–860.3. Fadini GP, Schiavon M, Cantini M, Baesso I, Facco M, Miorin M, et al. Circulating progenitor cells are reduced in patients with severe lung disease. Stem Cells. 2006. 24:1806–1813.4. Coraux C, Nawrocki-Raby B, Hinnrasky J, Kileztky C, Gaillard D, Dani C, et al. Embryonic stem cells generate airway epithelial tissue. Am J Respir Cell Mol Biol. 2005. 32:87–92.5. Wang G, Bunnell BA, Painter RG, Quiniones BC, Tom S, Lanson NA Jr, et al. Adult stem cells from bone marrow stroma differentiate into airway epithelial cells: potential therapy for cystic fibrosis. Proc Natl Acad Sci U S A. 2005. 102:186–191.6. Rojas M, Xu J, Woods CR, Mora AL, Spears W, Roman J, et al. Bone marrow-derived mesenchymal stem cells in repair of the injured lung. Am J Respir Cell Mol Biol. 2005. 33:145–152.7. Randell SH. Airway epithelial stem cells and the pathophysiology of chronic obstructive pulmonary disease. Proc Am Thorac Soc. 2006. 3:718–725.8. Al-Jamal R, Wallace WA, Harrison DJ. Gene therapy for chronic obstructive pulmonary disease: twilight or triumph? Expert Opin Biol Ther. 2005. 5:333–346.9. Ward C, Forrest IA, Murphy DM, Johnson GE, Robertson H, Cawston TE, et al. Phenotype of airway epithelial cells suggests epithelial to mesenchymal cell transition in clinically stable lung transplant recipients. Thorax. 2005. 60:865–871.10. Yamada M, Kubo H, Kobayashi S, Ishizawa K, Numasaki M, Ueda S, et al. Bone marrow-derived progenitor cells are important for lung repair after lipopolysaccharide-induced lung injury. J Immunol. 2004. 172:1266–1272.11. Tamama K, Fan VH, Griffith LG, Blair HC, Wells A. Epidermal growth factor as a candidate for ex vivo expansion of bone marrow-derived mesenchymal stem cells. Stem Cells. 2006. 24:686–695.12. Yen CC, Yang SH, Lin CY, Chen CM. Stem cells in the lung parenchyma and prospects for lung injury therapy. Eur J Clin Invest. 2006. 36:310–319.13. Kubo H. Is cell therapy in acute lung injury a realistic dream? Am J Respir Crit Care Med. 2005. 172:794–795.14. Kasai H, Allen JT, Mason RM, Kamimura T, Zhang Z. TGF-beta1 induces human alveolar epithelial to mesenchymal cell transition (EMT). Respir Res. 2005. 6:56.15. Stagg J. Immune regulation by mesenchymal stem cells: two sides to the coin. Tissue Antigens. 2007. 69:1–9.16. Uccelli A, Moretta L, Pistoia V. Immunoregulatory function of mesenchymal stem cells. Eur J Immunol. 2006. 36:2566–2573.17. Weiss DJ, Berberich MA, Borok Z, Gail DB, Kolls JK, Penland C, et al. Adult stem cells, lung biology, and lung disease. NHLBI/Cystic Fibrosis Foundation Workshop. Proc Am Thorac Soc. 2006. 3:193–207.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Clinical Applications of Neural Stem Cells for the Treatment of Peripheral Neuropathy
- Current Trends and Prospect of Cell Therapy using Hematopoietic Stem Cells
- Cell Biological Characteristics of Adult Stem Cells
- The Baiscs and the Prospect of Stem Cells for Pediatric Neurologic Diseases
- Stem Cells and Lung Regeneration
