Pioneering PGC-1αα–boosted secretome: a novel approach to combating liver fibrosis
- Affiliations
-
- 1Department of Surgery, Bucheon St. Mary’s Hospital, College of Medicine, the Catholic University of Korea, Seoul, Korea
- 2Department of Surgery, Seoul St. Mary’s Hospital, College of Medicine, the Catholic University of Korea, Seoul, Korea
- 3Catholic Central Laboratory of Surgery, College of Medicine, the Catholic University of Korea, Seoul, Korea
- 4Department of Surgery, Eunpyeong St. Mary’s Hospital, College of Medicine, the Catholic University of Korea, Seoul, Korea
- 5Translational Research Team, Surginex Co., Ltd., Seoul, Korea
- 6Department of Surgery, Daejeon St. Mary’s Hospital, College of Medicine, the Catholic University of Korea, Seoul, Korea
- 7Department of Surgery, Uijeongbu St. Mary’s Hospital, College of Medicine, the Catholic University of Korea, Seoul, Korea
- KMID: 2553385
- DOI: http://doi.org/10.4174/astr.2024.106.3.155
Abstract
- Purpose
Liver fibrosis is a critical health issue with limited treatment options. This study investigates the potential of PGC-Sec, a secretome derived from peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α)-overexpressing adipose-derived stem cells (ASCs), as a novel therapeutic strategy for liver fibrosis.
Methods
Upon achieving a cellular confluence of 70%–80%, ASCs were transfected with pcDNA-PGC-1α. PGC-Sec, obtained through concentration of conditioned media using ultrafiltration units with a 3-kDa cutoff, was assessed through in vitro assays and in vitro mouse models.
Results
In vitro, PGC-Sec significantly reduced LX2 human hepatic stellate cell proliferation and mitigated mitochondrial oxidative stress compared to the control-secretome. In an in vivo mouse model, PGC-Sec treatment led to notable reductions in hepatic enzyme activity, serum proinflammatory cytokine concentrations, and fibrosis-related marker expression. Histological analysis demonstrated improved liver histology and reduced fibrosis severity in PGC-Sec–treated mice. Immunohistochemical staining confirmed enhanced expression of PGC-1α, optic atrophy 1 (a mitochondrial function marker), and peroxisome proliferator-activated receptor alpha (an antifibrogenic marker) in the PGC-Sec–treated group, along with reduced collagen type 1A expression (a profibrogenic marker).
Conclusion
These findings highlight the therapeutic potential of PGC-Sec in combating liver fibrosis by enhancing mitochondrial biogenesis and function, and promoting antifibrotic processes. PGC-Sec holds promise as a novel treatment strategy for liver fibrosis.
Keyword
Figure
-

Fig. 1 Therapeutic effects of PGC-Sec on cell viability, exosome markers, and molecular markers in liver cells. (A) Schematic representation of PGC-1α–overexpressing ASCs generation by transfecting a plasmid encoding PGC-1α. (B) Comparison of CD81 expression, an exosome marker, in the secretome obtained from PGC-Sec and Ctrl-Sec. PGC-Sec exhibited a 3.32-fold higher secretion of CD81 compared to Ctrl-Sec. (C) ELISA showing the secretion of SOD mRNA in PGC-Sec and Ctrl-Sec. PGC-Sec displayed a 3.43-fold higher expression of SOD mRNA compared to Ctrl-Sec. (D) Viability of LX2 cells (human hepatic stellate cells) and AML12 cells (mouse hepatocytes) treated with Ctrl-Sec or PGC-Sec, with or without (TAA) treatment, as assessed by cell viability assay. PGC-Sec treatment resulted in the most significant decrease in TAA-treated LX2 cell viability and the most substantial increase in TAA-treated AML12 cell viability. (E) Effects of Ctrl-Sec and PGC-Sec on apoptosis and proliferation in AML12 cells. Expression of the antiapoptotic marker Bcl-xL and the proliferation marker PCNA decreased in AML12 cells upon TAA treatment. Treatment with Ctrl-Sec increased the expression of these markers, while treatment with PGC-Sec showed the most significant increase. Values are presented as mean ± standard deviation of 3 independent experiments. PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; PGC-Sec, PGC-1α-boosted secretome; ASC, adipose-derived stem cell; SOD, superoxide dismutase; mRNA, messenger RNA; Ctrl-Sec, control-secretome; TAA, thioacetamide; MCL-1, myeloid cell leukemia 1; PCNA, proliferating cell nuclear antigen. *P < 0.05.
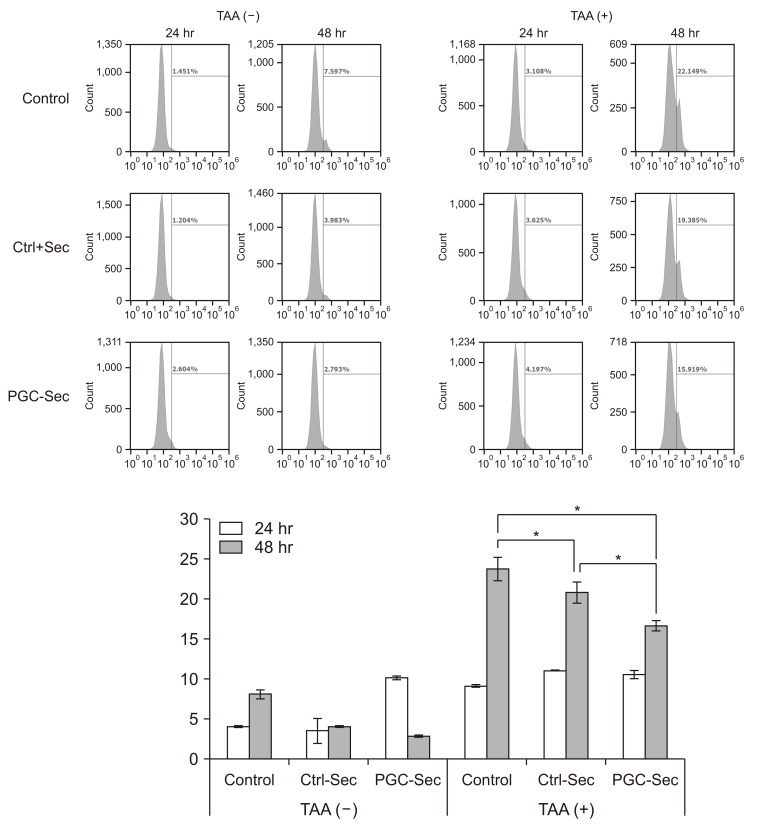
Fig. 2 Determination of effects of PGC-Sec on mitochondrial reactive oxygen species production. Mitochondrial superoxide indicator (MitoSOX) flow cytometry analysis of mitochondrial superoxide production in control AML12 cells and TAA-induced liver injury cells, indicating a significant reduction in MitoSOX fluorescence in both Ctrl-Sec and PGC-Sec treatment groups compared to the no treatment group, with a more pronounced reduction in the PGC-Sec treatment group. Values are presented as mean ± standard deviation of 3 independent experiments. Ctrl-Sec, Control-secretome, PGC-Sec, PGC-1α-boosted secretome; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; TAA, thioacetamide; MitoSOX, mitochondrial superoxide indicator. *P < 0.05.
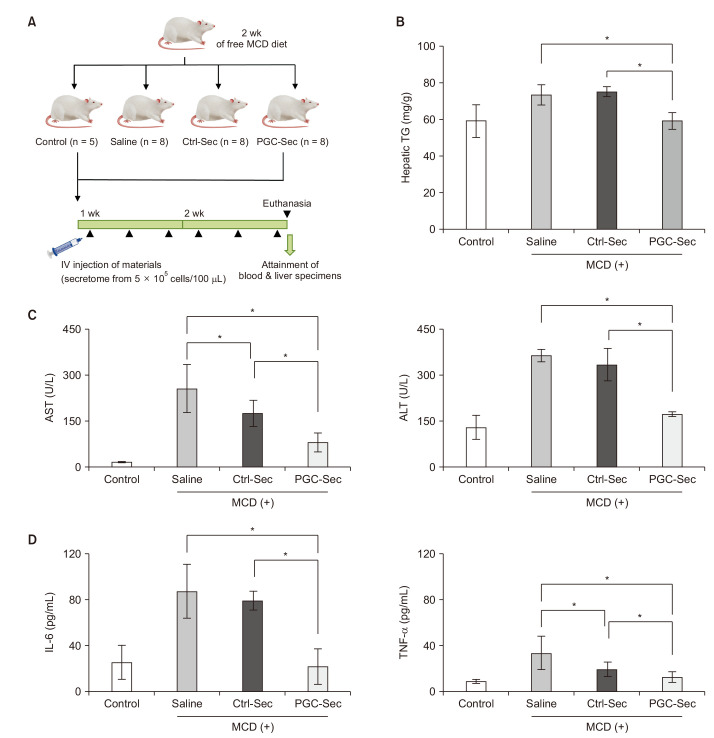
Fig. 3 In vitro effects of PGC-Sec on liver fibrosis; reduction in TG levels, liver enzymes, and proinflammatory cytokines. (A) Animal experimental design. (B) Assessment of TG levels in liver tissue samples from control mice, saline-injected mice, Ctrl-Sec–injected mice, and PGC-Sec–injected mice. Treatment with PGC-Sec significantly reduced TG levels, restoring them to levels comparable to the control group. (C) Measurement of serum liver enzyme levels, including AST and ALT, in the experimental groups. The PGC-Sec group showed significantly lower liver enzyme levels compared to the Ctrl-Sec group. (D) Evaluation of serum levels of proinflammatory cytokines, IL-6, and TNF-α in the experimental groups. The PGC-Sec group exhibited a significantly greater reduction in IL-6 and TNF-α levels compared to the Ctrl-Sec group. Values are presented as mean ± standard deviation of 3 independent experiments. IV, intravenous; MCD, methionine/choline-deficient; Ctrl-Sec, control-secretome; PGC-Sec, PGC-1α-boosted secretome; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha. *P < 0.05.
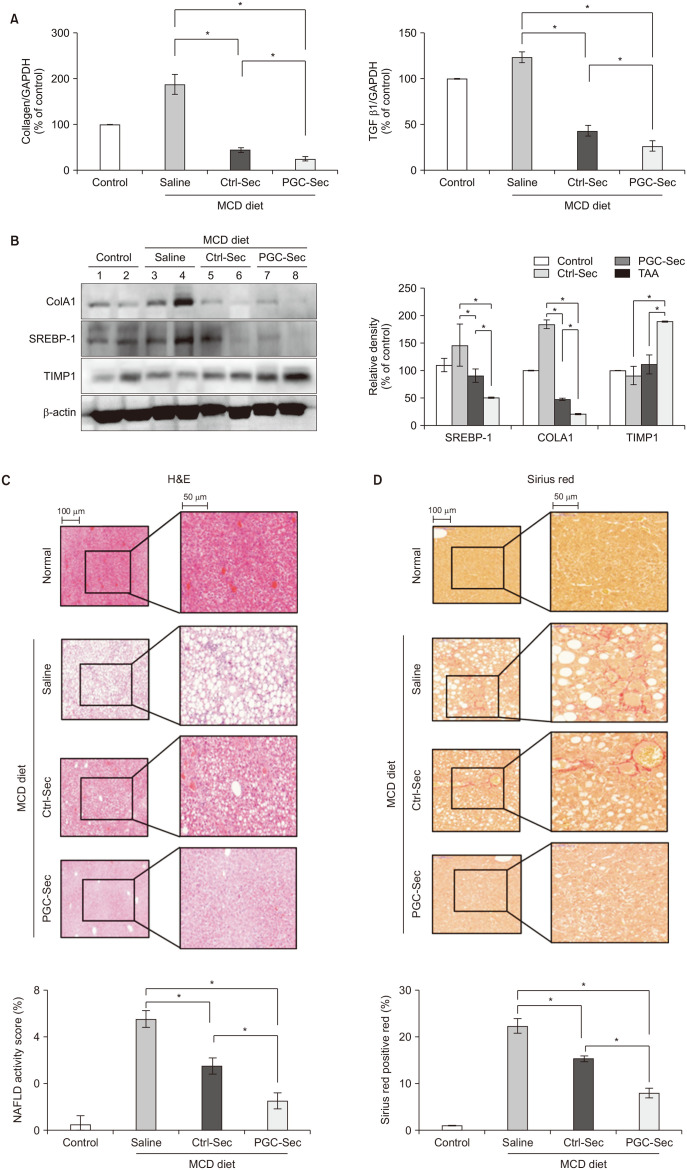
Fig. 4 In vivo effects of PGC-Sec on liver fibrosis; protein expression and histological improvement. (A) Quantitative real-time PCR analysis of liver fibrosis-associated messenger RNA (mRNA) expression, including collagen and TGF-β1, in liver specimens. Both secretome injection groups exhibited significantly reduced expression of collagen and TGF-β mRNA compared to the saline injection group. The PGC-Sec group displayed a more pronounced decrease in collagen and TGF-β1 mRNA expression compared to the Ctrl-Sec group. (B) Western blot analysis of protein levels of ColA1, TIMP1, and SREBP-1 in liver specimens. Secretome injection groups showed significantly lower ColA1 levels and higher TIMP levels compared to the saline injection group. The PGC-Sec group exhibited significantly increased TIMP expression compared to the Ctrl-Sec group. Both Ctrl-Sec and PGC-Sec displayed a decrease in SREBP-1 levels, with PGC-Sec showing the most significant reduction. (C) Histological analysis of liver fibrosis severity using H&E staining. The secretome injection groups demonstrated a significant reduction in liver fibrosis severity compared to the saline injection group, with the PGC-Sec group showing the most substantial improvement. (D) Evaluation of liver fibrosis using Sirius red staining, a reliable method for visualizing and assessing liver fibrosis. Both Ctrl-Sec and PGC-Sec groups exhibited a significant reduction in liver fibrosis compared to the saline injection group. Values are presented as mean ± standard deviation of 3 independent experiments. PGC-Sec, PGC-1α-boosted secretome; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Ctrl-Sec, control-secretome; MCD, methionine/choline-deficient; ColA1, collagen type A1; SREBP, sterol regulatory element-binding protein; TIMP, tissue inhibitor of metalloproteinases; TAA, thioacetamide; SREBP-1, sterol regulatory element-binding protein 1; NAFLD, non-alcoholic fatty liver disease. *P < 0.05.

Fig. 5 Immunohistochemistry validating the antifibrogenic effects of PGC-Sec in a mouse model of liver fibrosis. (A) PGC-1α immunohistochemistry showing significantly elevated PGC-1α expression in the PGC-Sec group. (B) OPA1 expression comparison across different groups, with the highest OPA1 expression observed in the PGC-Sec group, indicating enhanced mitochondrial function. (C) ColA1 immunohistochemistry revealing markedly reduced expression of the profibrogenic marker in the PGC-Sec group. (D) PPAR-α immunohistochemistry showing significantly increased expression of the antifibrogenic marker in the PGC-Sec group. These findings emphasize the profound impact of PGC-Sec treatment on enhancing mitochondrial biogenesis and function, as well as promoting antifibrosis processes, collectively highlighting its potential as a novel therapeutic strategy for liver fibrosis. Percentages of immunoreactive areas were measured using ImageJ of the National Institutes of Health and expressed as relative values to those in normal livers. MCD, methionine/choline-deficient; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; PGC-Sec, PGC-1α-boosted secretome; Ctrl-Sec, control-secretome; OPA1, optic atrophy 1; ColA1, collagen type A1; PPAR-α, peroxisome proliferator-activated receptor alpha. *P < 0.05.
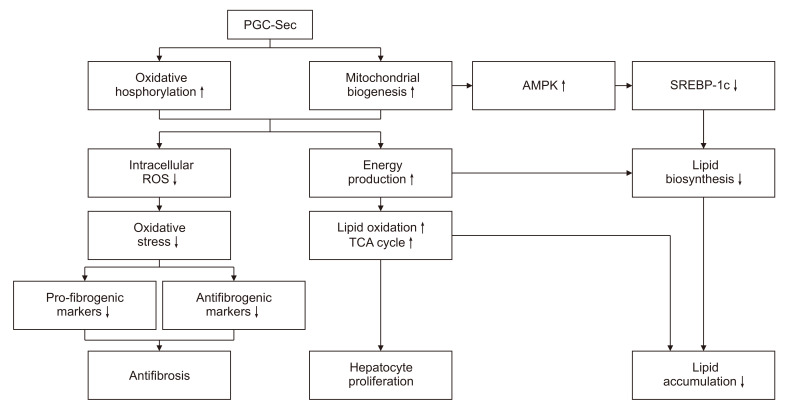
Fig. 6 The postulated mechanism by which PGC-Sec contributes to antifibrosis. PGC-Sec activates mitochondrial biogenesis and oxidative phosphorylation, leading to a decrease in intracellular ROS and oxidative stress. This reduction in oxidative stress results in decreased expression of profibrogenic markers and increased expression of antifibrosis markers, thereby promoting antifibrosis. PGC-Sec also enhances mitochondrial function, leading to increased energy production, which in turn promotes the proliferation of liver cells. Furthermore, it has been reported that increased mitochondrial biogenesis by PGC-Sec results in increased AMPK activation. This, as evidenced by our experimental results, leads to a decrease in SREBP-1c, a key regulator of lipid biogenesis, and consequently a reduction in lipid accumulation within hepatocytes. PGC-Sec, PGC-1α-boosted secretome; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; ROS, reactive oxygen species; AMPK, AMP-activated protein kinase; SREBP, sterol regulatory element-binding protein; TCA, tricarboxylic acid.
Reference
-
1. Pellicoro A, Ramachandran P, Iredale JP, Fallowfield JA. Liver fibrosis and repair: immune regulation of wound healing in a solid organ. Nat Rev Immunol. 2014; 14:181–194. PMID: 24566915.2. Geissler S, Textor M, Kühnisch J, Könnig D, Klein O, Ode A, et al. Functional comparison of chronological and in vitro aging: differential role of the cytoskeleton and mitochondria in mesenchymal stromal cells. PLoS One. 2012; 7:e52700. PMID: 23285157.3. Phinney DG, Pittenger MF. Concise review: MSC-derived exosomes for cell-free therapy. Stem Cells. 2017; 35:851–858. PMID: 28294454.4. Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014; 24:R453–R462. PMID: 24845678.5. Sies H, Jones DP. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat Rev Mol Cell Biol. 2020; 21:363–383. PMID: 32231263.6. Austin S, St-Pierre J. PGC1α and mitochondrial metabolism--emerging concepts and relevance in ageing and neurodegenerative disorders. J Cell Sci. 2012; 125:4963–4971. PMID: 23277535.7. LeBleu VS, O'Connell JT, Gonzalez Herrera KN, Wikman H, Pantel K, Haigis MC, et al. PGC-1α mediates mitochondrial biogenesis and oxidative phosphorylation in cancer cells to promote metastasis. Nat Cell Biol. 2014; 16:992–1003. 1–15. PMID: 25241037.8. Lee SC, Jeong HJ, Lee SK, Kim SJ. Hypoxic conditioned medium from human adipose-derived stem cells promotes mouse liver regeneration through JAK/STAT3 signaling. Stem Cells Transl Med. 2016; 5:816–825. PMID: 27102647.9. Kim OH, Hong HE, Seo H, Kwak BJ, Choi HJ, Kim KH, et al. Generation of induced secretome from adipose-derived stem cells specialized for disease-specific treatment: an experimental mouse model. World J Stem Cells. 2020; 12:70–86. PMID: 32110276.10. Paik KY, Kim KH, Park JH, Lee JI, Kim OH, Hong HE, et al. A novel antifibrotic strategy utilizing conditioned media obtained from miR-150-transfected adipose-derived stem cells: validation of an animal model of liver fibrosis. Exp Mol Med. 2020; 52:438–449. PMID: 32152450.11. Kim JG, Lee SC, Kim OH, Kim KH, Song KY, Lee SK, et al. HSP90 inhibitor 17-DMAG exerts anticancer effects against gastric cancer cells principally by altering oxidant-antioxidant balance. Oncotarget. 2017; 8:56473–56489. PMID: 28915605.12. Scarpulla RC. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim Biophys Acta. 2011; 1813:1269–1278. PMID: 20933024.13. Summermatter S, Santos G, Pérez-Schindler J, Handschin C. Skeletal muscle PGC-1α controls whole-body lactate homeostasis through estrogen-related receptor α-dependent activation of LDH B and repression of LDH A. Proc Natl Acad Sci U S A. 2013; 110:8738–8743. PMID: 23650363.14. Lai L, Leone TC, Zechner C, Schaeffer PJ, Kelly SM, Flanagan DP, et al. Transcriptional coactivators PGC-1alpha and PGC-lbeta control overlapping programs required for perinatal maturation of the heart. Genes Dev. 2008; 22:1948–1961. PMID: 18628400.15. Zhang Q, Chen W, Xie C, Dai X, Ma J, Lu J. The role of PGC-1α in digestive system malignant tumours. Anticancer Agents Med Chem. 2020; 20:276–285. PMID: 31702508.16. Zhang Y, Shen L, Zhu H, Dreissigacker K, Distler D, Zhou X, et al. PGC-1α regulates autophagy to promote fibroblast activation and tissue fibrosis. Ann Rheum Dis. 2020; 79:1227–1233. PMID: 32482644.17. Wu L, Mo W, Feng J, Li J, Yu Q, Li S, et al. Astaxanthin attenuates hepatic damage and mitochondrial dysfunction in non-alcoholic fatty liver disease by up-regulating the FGF21/PGC-1α pathway. Br J Pharmacol. 2020; 177:3760–3777. PMID: 32446270.18. Lee G, Uddin MJ, Kim Y, Ko M, Yu I, Ha H. PGC-1α, a potential therapeutic target against kidney aging. Aging Cell. 2019; 18:e12994. PMID: 31313501.19. Han Y, Xu X, Tang C, Gao P, Chen X, Xiong X, et al. Reactive oxygen species promote tubular injury in diabetic nephropathy: the role of the mitochondrial ros-txnip-nlrp3 biological axis. Redox Biol. 2018; 16:32–46. PMID: 29475133.20. Cheng CF, Ku HC, Lin H. PGC-1α as a pivotal factor in lipid and metabolic regulation. Int J Mol Sci. 2018; 19:3447. PMID: 30400212.21. Baglio SR, Rooijers K, Koppers-Lalic D, Verweij FJ, Pérez Lanzón M, Zini N, et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res Ther. 2015; 6:127. PMID: 26129847.22. Rani S, Ryan AE, Griffin MD, Ritter T. Mesenchymal stem cell-derived extracellular vesicles: toward cell-free therapeutic applications. Mol Ther. 2015; 23:812–823. PMID: 25868399.23. Jäger S, Handschin C, St-Pierre J, Spiegelman BM. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc Natl Acad Sci U S A. 2007; 104:12017–12022. PMID: 17609368.24. Suwa M, Egashira T, Nakano H, Sasaki H, Kumagai S. Metformin increases the PGC-1alpha protein and oxidative enzyme activities possibly via AMPK phosphorylation in skeletal muscle in vivo. J Appl Physiol (1985). 2006; 101:1685–1692. PMID: 16902066.25. Yap F, Craddock L, Yang J. Mechanism of AMPK suppression of LXR-dependent Srebp-1c transcription. Int J Biol Sci. 2011; 7:645–650. PMID: 21647332.26. Li W, Li Y, Wang Q, Yang Y. Crude extracts from Lycium barbarum suppress SREBP-1c expression and prevent diet-induced fatty liver through AMPK activation. Biomed Res Int. 2014; 2014:196198. PMID: 25013763.27. Han Y, Hu Z, Cui A, Liu Z, Ma F, Xue Y, et al. Post-translational regulation of lipogenesis via AMPK-dependent phosphorylation of insulin-induced gene. Nat Commun. 2019; 10:623. PMID: 30733434.28. Abu Shelbayeh O, Arroum T, Morris S, Busch KB. PGC-1α is a master regulator of mitochondrial lifecycle and ROS stress response. Antioxidants (Basel). 2023; 12:1075. PMID: 37237941.29. Halling JF, Pilegaard H. PGC-1α-mediated regulation of mitochondrial function and physiological implications. Appl Physiol Nutr Metab. 2020; 45:927–936. PMID: 32516539.30. Estornut C, Milara J, Bayarri MA, Belhadj N, Cortijo J. Targeting oxidative stress as a therapeutic approach for idiopathic pulmonary fibrosis. Front Pharmacol. 2021; 12:794997. PMID: 35126133.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Isolation of Secretome with Enhanced Antifibrotic Properties from miR-214-Transfected Adipose-Derived Stem Cells
- PGC1-alpha plays a role in hypothermic renal protection of renal fibrosis after acute kidney injury
- Hypothermia protects against renal fibrosis after ischemia reperfusion injury
- A novel strategy to promote liver regeneration: utilization of secretome obtained from survivin-overexpressing adipose-derived stem cells
- Effect of Ritonavir-boosting on Atazanavir Discontinuation due to Jaundice in HIV-infected Koreans
