FoxO6-Mediated TXNIP Induces Lipid Accumulation in the Liver through NLRP3 Inflammasome Activation
- Affiliations
-
- 1Department of Life Sciences, Chosun University College of Natural Science, Gwangju, Korea
- 2Department of Pharmacy, Kyungsung University College of Pharmacy, Busan, Korea
- 3Department of Food Science & Technology, Pusan National University College of Natural Resources and Life Science, Miryang, Korea
- KMID: 2552804
- DOI: http://doi.org/10.3803/EnM.2023.1826
Abstract
- Background
Hepatic steatosis, which involves the excessive accumulation of lipid droplets in hepatocytes, presents a significant global health concern due to its association with obesity and metabolic disorders. Inflammation plays a crucial role in the progression of hepatic steatosis; however, the precise molecular mechanisms responsible for this process remain unknown.
Methods
This study investigated the involvement of the nucleotide-binding oligomerization domain-like receptor pyrin domain-containing-3 (NLRP3) inflammasome and the forkhead box O6 (FoxO6) transcription factor in the pathogenesis of hepatic steatosis. We monitored the NLRP3 inflammasome and lipogenesis in mice overexpressing the constitutively active (CA)-FoxO6 allele and FoxO6-null mice. In an in vitro study, we administered palmitate to liver cells overexpressing CA-FoxO6 and measured changes in lipid metabolism.
Results
We administered palmitate treatment to clarify the mechanisms through which FoxO6 activates cytokine interleukin (IL)-1β through the NLRP3 inflammasome. The initial experiments revealed that dephosphorylation led to palmitate-induced FoxO6 transcriptional activity. Further palmitate experiments showed increased expression of IL-1β and the hepatic NLRP3 inflammasome complex, including adaptor protein apoptotic speck protein containing a caspase recruitment domain (ASC) and pro-caspase-1. Furthermore, thioredoxin-interacting protein (TXNIP), a key regulator of cellular redox conditions upstream of the NLRP3 inflammasome, was induced by FoxO6 in the liver and HepG2 cells.
Conclusion
The findings of this study shed light on the molecular mechanisms underpinning the FoxO6-NLRP3 inflammasome axis in promoting inflammation and lipid accumulation in the liver.
Keyword
Figure
-
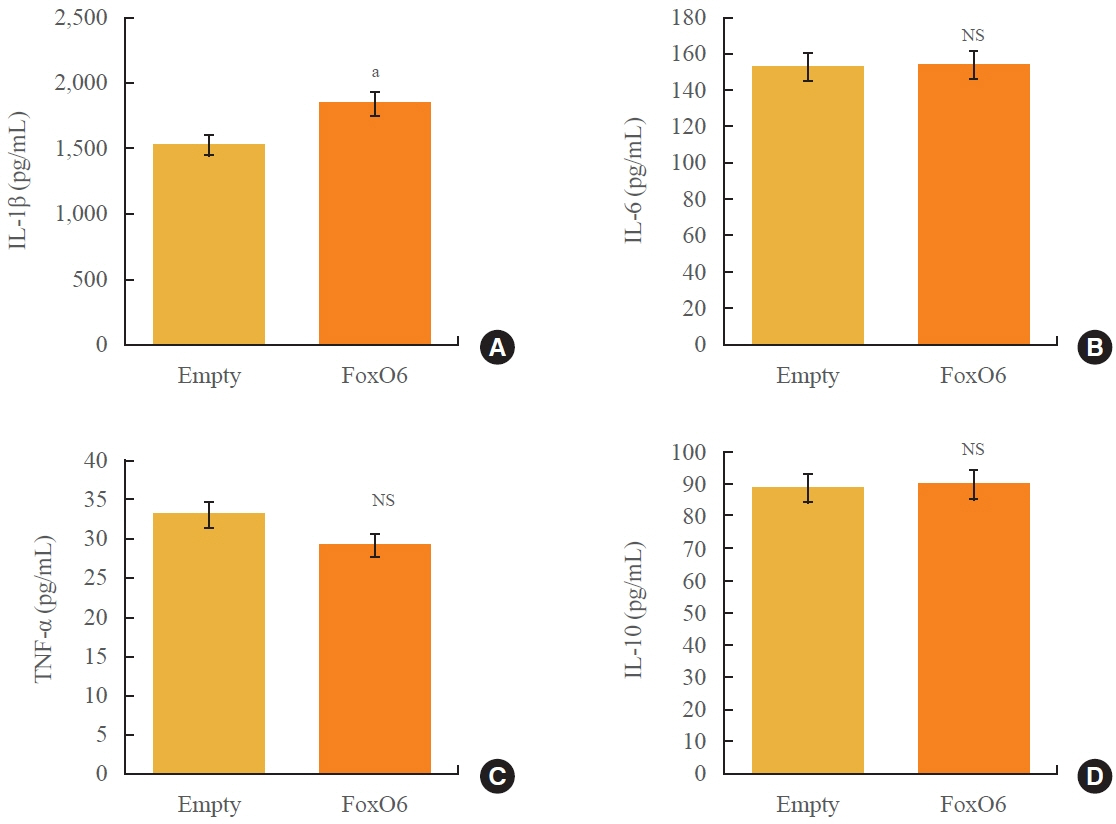
Fig. 1. Effects of forkhead box O6 (FoxO6) on cytokine production. Serum levels of pro-cytokines in mice injected with either constitutively active form of FoxO6 (FoxO6-CA) or null adenoviral vectors, assessed over a period of 13 days: enzyme-linked immunosorbent assay (ELISA) of (A) interleukin (IL)-1β, (B) IL-6, (C) tumor necrosis factor α (TNF-α), and (D) IL-10 (n=5 in each group). Three experiments were run for each protein, which yielded similar results; a single representative result is shown here for each protein. NS, not significant. aP<0.01 vs. the results obtained using the empty adenovirus vector by paired t test.
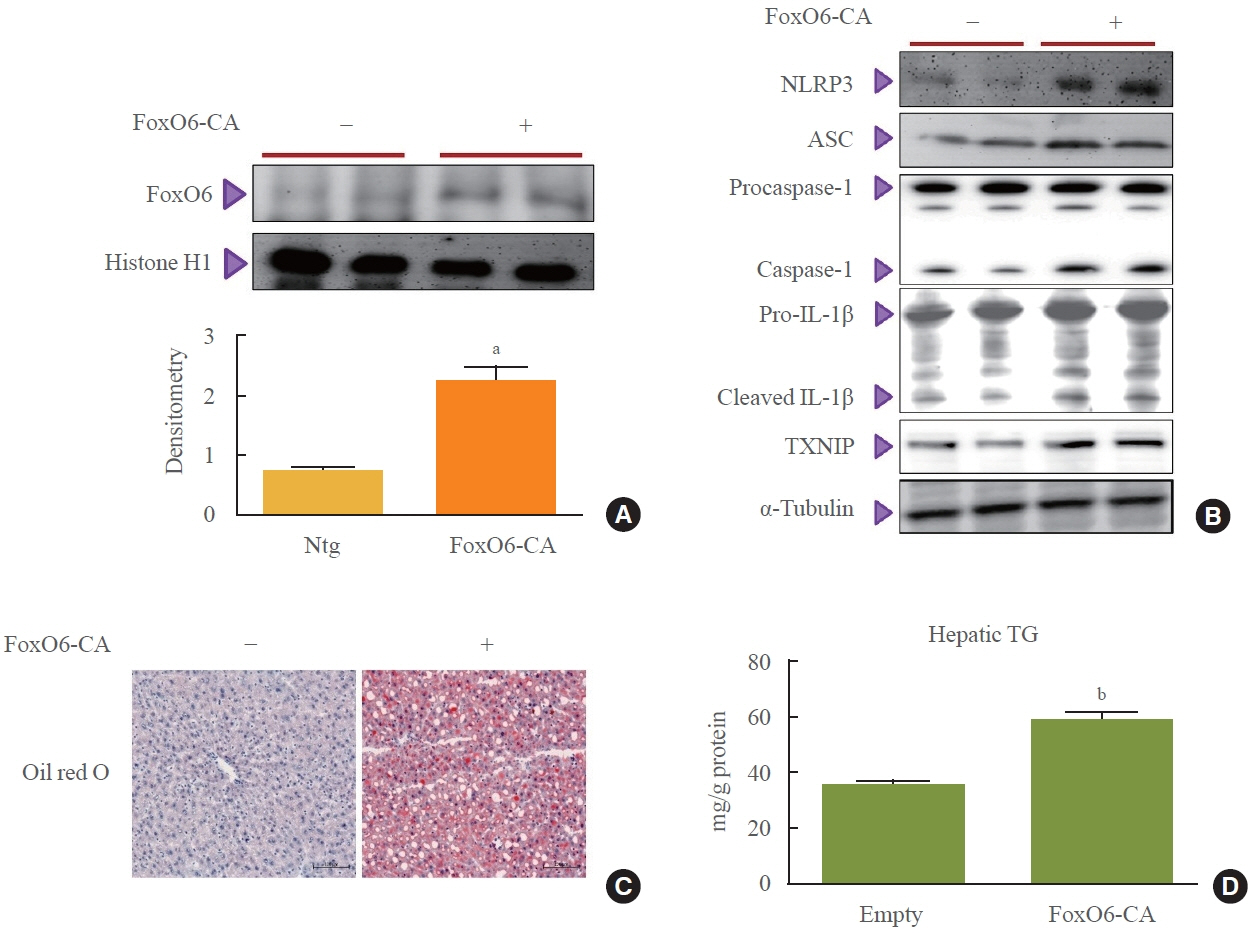
Fig. 2. Forkhead box O6 (FoxO6) regulates nucleotide-binding oligomerization domain-like receptor pyrin domain-containing-3 (NLRP3) inflammasome and lipid accumulation in the liver of mice injected with an adenoviral vector expressing FoxO6. (A) Western blotting was used to detect FoxO6 in the nuclear extracts (20 μg of protein) of liver tissues. Bars in densitometry data represent mean±standard error, and significance was determined using the unpaired test non-trangenic (Ntg). (B) FoxO6 activated the NLRP3 inflammasome genes. Western blotting was used to detect NLRP3, apoptotic speck protein containing a caspase recruitment domain (ASC), caspase-1, interleukin (IL)-1β, and thioredoxin-interacting protein (TXNIP) in cytoplasmic extracts (20 μg of protein) from the liver. (C) Livers were stained with Oil Red O to visualize lipid accumulation (bar=100 μm). (D) Triglyceride levels in hepatic tissues. FoxO6-CA, constitutively active form of FoxO6. aP<0.01 vs. non-transgenic (n=3 each); bP<0.001 vs. empty vector virus by paired t test.
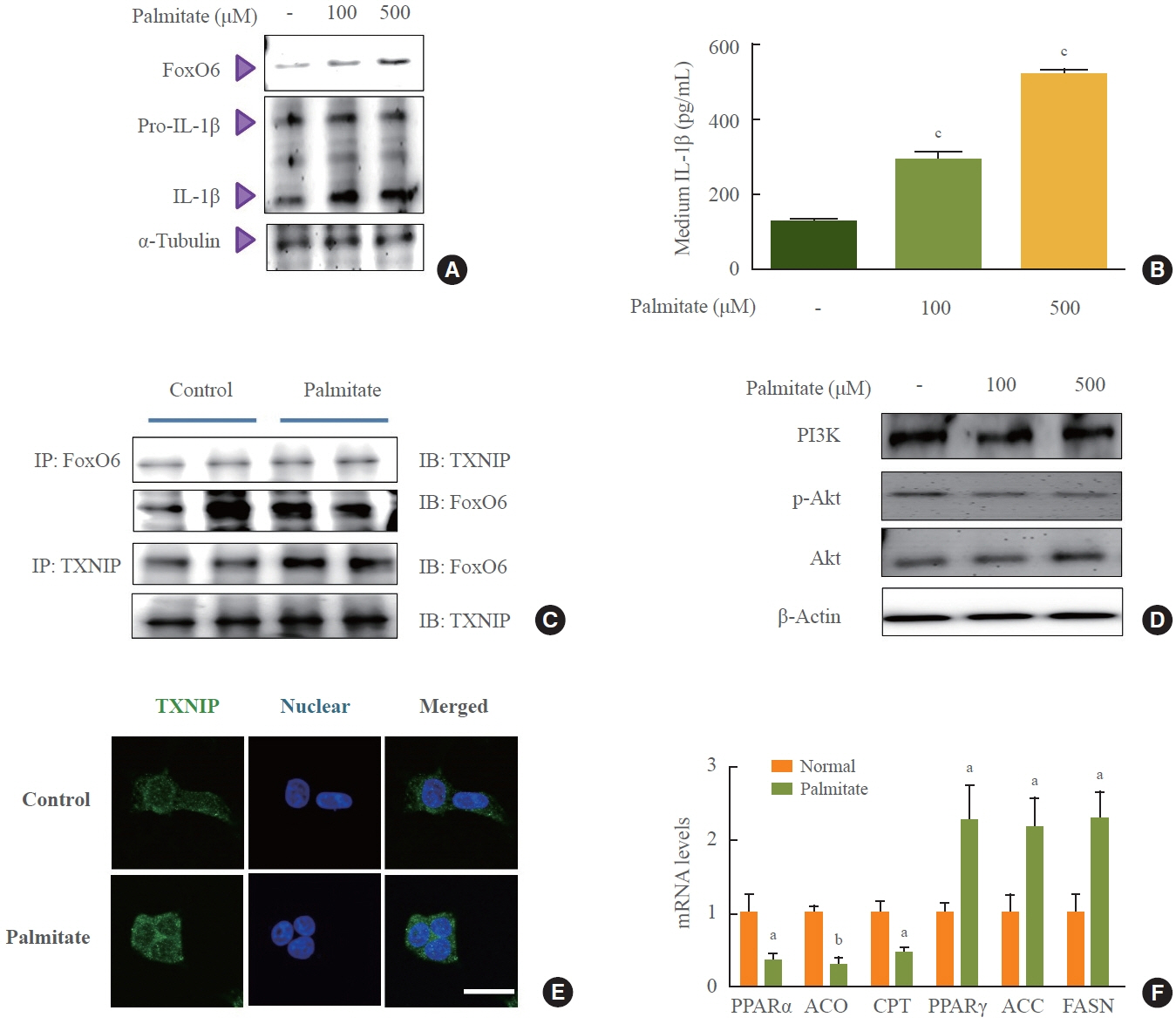
Fig. 3. Interaction between forkhead box O6 (FoxO6) and thioredoxin-interacting protein (TXNIP) induced interleukin (IL)-1β in palmitatetreated HepG2 cells. HepG2 cells were treated with palmitate at various concentrations, and FoxO6 levels were determined by Western blotting. Samples loaded on gels were probed with α-tubulin. (A) Levels of FoxO6 and IL-1β noticeably increased after treatment with 100 and 500 μM palmitate. (B) Effects of palmitate on IL-1β production. The conditioned medium from the experiment in (A) was used for the determination of IL-1β levels. (C) Western blotting showed that immunoprecipitated FoxO6 and TXNIP were physically associated with each other. (D) Levels of phosphoinositide 3-kinase (PI3K), phosphorylated protein kinase B (p-Akt), and total-Akt were noticeably diminished after treatment with 100 and 500 μM palmitate. (E) HepG2 cells were treated with palmitate (500 μM) for 24 hours. Cells were immunostained using rabbit anti-TXNIP antibody followed by immunoglobulin G conjugated with fluorescein isothiocyanate (green) (bar=50 μm). (F) β-Oxidation and lipogenesis genes were subjected to real-time quantitative reverse-transcription polymerase chain reaction analysis. IP, immunoprecipitation; IB, immunoblotting; PPARα, peroxisome proliferator-activated receptor alpha; ACO, acyl-coA oxidase; CPT, carnitine palmitoyltransferase; PPARγ, peroxisome proliferator-activated receptor gamma; ACC, acetyl-coA carboxylase; FASN, fatty acid synthase. aP<0.05 and bP< 0.01 vs. normal by paired t test; cP<0.001 vs. vehicle by one-way analysis of variance (ANOVA) with Tukey’s post hoc test.
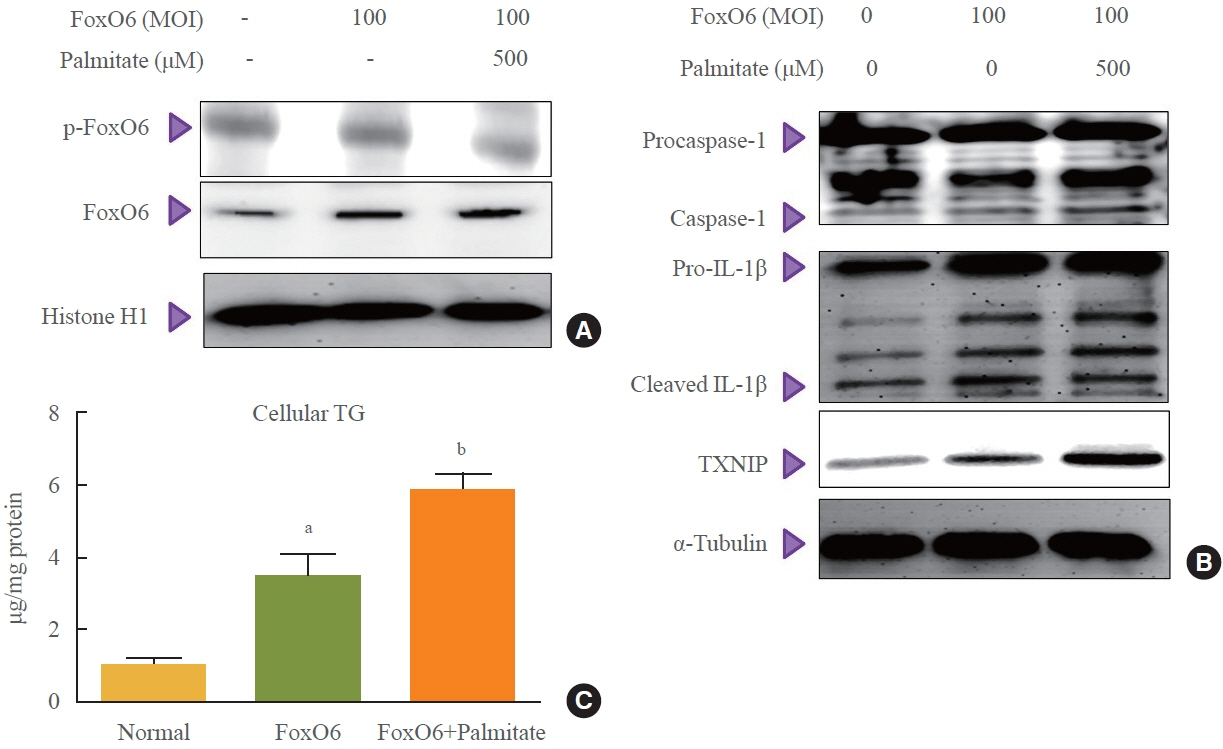
Fig. 4. Palmitate-induced forkhead box O6 (FoxO6) regulated the nucleotide-binding oligomerization domain-like receptor pyrin domaincontaining- 3 (NLRP3) inflammasome in HepG2 cells. (A) Phosphorylated FoxO6 (p-FoxO6) and total-FoxO6 expression in nuclear. (B) Cytosolic levels of thioredoxin-interacting protein (TXNIP), caspase-1, and interleukin (IL)-1β noticeably increased in FoxO6-transfected (100 multiplicity of infection [MOI]) cells after treatment with 500 μM palmitate. (C) Triglyceride levels in liver cells. aP<0.01 vs. empty vector virus; bP<0.05 vs. FoxO6 virus by one-way analysis of variance (ANOVA) with Tukey’s post hoc test.
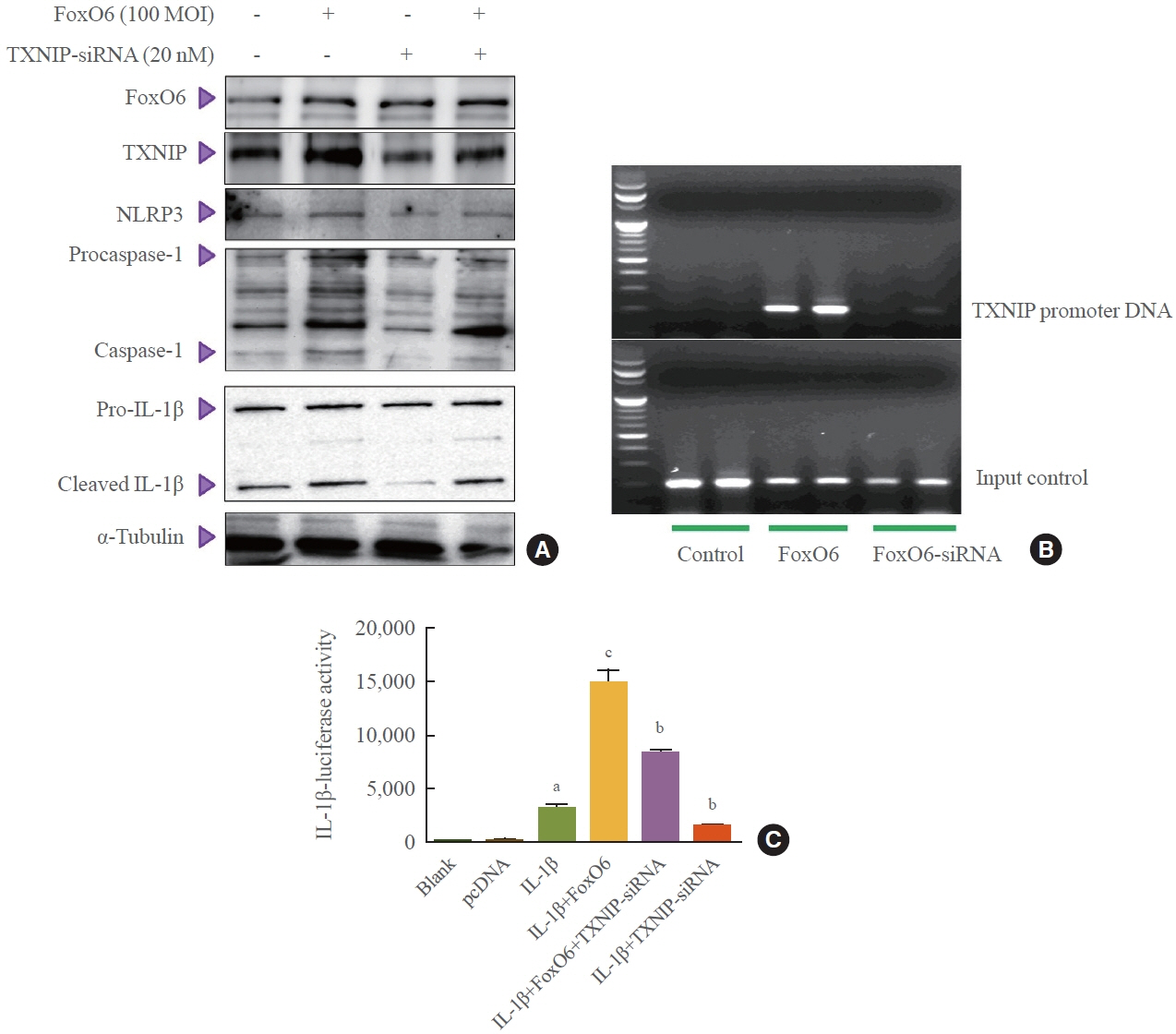
Fig. 5. Forkhead box O6 (FoxO6)-induced thioredoxin-interacting protein (TXNIP) regulated the nucleotide-binding oligomerization domain-like receptor pyrin domain-containing-3 (NLRP3) inflammasome. (A) Pre-treated (1 day) with or without TXNIP-small interfering RNA (siRNA; 20 nM), and then stimulated with FoxO6 (100 multiplicity of infection [MOI]) in cells, and analyzed by Western blotting. (B) FoxO6 bound to the TXNIP promoter in HepG2 cells transfected with a vector containing FoxO6 (100 MOI) or FoxO6-siRNA (100 MOI). After incubation for 24 hours, the cells were subjected to a chromatin immunoprecipitation (ChIP) assay using rabbit pre-immune immunoglobulin G (lanes 1, 2) or anti-FoxO6 antibody (lanes 3–6). The immunoprecipitates were subjected to polymerase chain reaction using the TXNIP promoter. (C) Effect of wild-type FoxO6 with or without TXNIP-siRNA (20 nM) on the activity of the interleukin (IL)-1β promoter. HepG2 cells in 48-well microplates were transduced with adenoviral vector (AdV)-FoxO6, TXNIP-siRNA (20 nM), or control AdV-null vectors at a fixed dose (MOI, 100 pfu/cell), followed by transfection with 0.5 μg of pcDNA and IL-1β DNA in the culture medium. After incubation for 24 hours, the cells were harvested and treated with palmitate (500 μM); luciferase and β-galactosidase activities were determined 8 hours after palmitate treatment. The relative luciferase activity was calculated based on the IL-1β-luciferase/β-galactosidase activity ratio. The data are expressed as mean±standard error of the mean. aP<0.001 vs. pcDNA treated cells; bP<0.01, cP<0.001 vs. IL-1β DNA treated cells by one-way analysis of variance (ANOVA) with Tukey’s post hoc test.

Fig. 6. Changes in the nucleotide-binding oligomerization domain-like receptor pyrin domain-containing-3 (NLRP3) inflammasome in response to forkhead box O6 (FoxO6) depletion. (A) Western blot was used to detect FoxO6, thioredoxin-interacting protein (TXNIP), NLRP3, apoptotic speck protein containing a caspase recruitment domain (ASC), caspase-1, and interleukin (IL)-1β in liver tissues; β-actin levels were used as loading controls. (B) The expression of relevant genes (FoxO6, insulin receptor substrate 1 [IRS1], IRS2, and IL-1β) was analyzed by quantitative polymerase chain reaction. The results were normalized with respect to actin levels. (C) Possible mechanism by which FoxO6 activates the NLRP3 inflammasome in insulin resistance. WT, wild-type; KO, knockout; TG, triglyceride; p, phospho; Ser, serine; Tyr, tyrosine; Akt, protein kinase B. aP<0.001 vs. WT by paired t test.
Reference
-
1. Van Der Heide LP, Hoekman MF, Smidt MP. The ins and outs of FoxO shuttling: mechanisms of FoxO translocation and transcriptional regulation. Biochem J. 2004; 380(Pt 2):297–309.2. Accili D, Arden KC. FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell. 2004; 117:421–6.3. Barthel A, Schmoll D, Unterman TG. FoxO proteins in insulin action and metabolism. Trends Endocrinol Metab. 2005; 16:183–9.4. Biggs WH 3rd, Meisenhelder J, Hunter T, Cavenee WK, Arden KC. Protein kinase B/Akt-mediated phosphorylation promotes nuclear exclusion of the winged helix transcription factor FKHR1. Proc Natl Acad Sci U S A. 1999; 96:7421–6.5. Kawamori D, Kaneto H, Nakatani Y, Matsuoka TA, Matsuhisa M, Hori M, et al. The forkhead transcription factor Foxo1 bridges the JNK pathway and the transcription factor PDX-1 through its intracellular translocation. J Biol Chem. 2006; 281:1091–8.6. Martinez SC, Tanabe K, Cras-Meneur C, Abumrad NA, Bernal-Mizrachi E, Permutt MA. Inhibition of Foxo1 protects pancreatic islet beta-cells against fatty acid and endoplasmic reticulum stress-induced apoptosis. Diabetes. 2008; 57:846–59.7. Hotamisligil GS. Inflammation and metabolic disorders. Nature. 2006; 444:860–7.8. Schenk S, Saberi M, Olefsky JM. Insulin sensitivity: modulation by nutrients and inflammation. J Clin Invest. 2008; 118:2992–3002.9. Kamagate A, Dong HH. FoxO1 integrates insulin signaling to VLDL production. Cell Cycle. 2008; 7:3162–70.10. Kim DH, Lee B, Lee J, Kim ME, Lee JS, Chung JH, et al. FoxO6-mediated IL-1β induces hepatic insulin resistance and age-related inflammation via the TF/PAR2 pathway in aging and diabetic mice. Redox Biol. 2019; 24:101184.11. Kim DH, Perdomo G, Zhang T, Slusher S, Lee S, Phillips BE, et al. FoxO6 integrates insulin signaling with gluconeogenesis in the liver. Diabetes. 2011; 60:2763–74.12. Vandanmagsar B, Youm YH, Ravussin A, Galgani JE, Stadler K, Mynatt RL, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat Med. 2011; 17:179–88.13. Donath MY. Targeting inflammation in the treatment of type 2 diabetes: time to start. Nat Rev Drug Discov. 2014; 13:465–76.14. Schroder K, Tschopp J. The inflammasomes. Cell. 2010; 140:821–32.15. Febbraio MA. Role of interleukins in obesity: implications for metabolic disease. Trends Endocrinol Metab. 2014; 25:312–9.16. Hoque R, Vodovotz Y, Mehal W. Therapeutic strategies in inflammasome mediated diseases of the liver. J Hepatol. 2013; 58:1047–52.17. Chen J, Saxena G, Mungrue IN, Lusis AJ, Shalev A. Thioredoxin-interacting protein: a critical link between glucose toxicity and beta-cell apoptosis. Diabetes. 2008; 57:938–44.18. Perrone L, Devi TS, Hosoya K, Terasaki T, Singh LP. Thioredoxin interacting protein (TXNIP) induces inflammation through chromatin modification in retinal capillary endothelial cells under diabetic conditions. J Cell Physiol. 2009; 221:262–72.19. Zhou R, Tardivel A, Thorens B, Choi I, Tschopp J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol. 2010; 11:136–40.20. Minn AH, Hafele C, Shalev A. Thioredoxin-interacting protein is stimulated by glucose through a carbohydrate response element and induces beta-cell apoptosis. Endocrinology. 2005; 146:2397–405.21. Parikh H, Carlsson E, Chutkow WA, Johansson LE, Storgaard H, Poulsen P, et al. TXNIP regulates peripheral glucose metabolism in humans. PLoS Med. 2007; 4:e158.22. Li X, Rong Y, Zhang M, Wang XL, LeMaire SA, Coselli JS, et al. Up-regulation of thioredoxin interacting protein (Txnip) by p38 MAPK and FOXO1 contributes to the impaired thioredoxin activity and increased ROS in glucose-treated endothelial cells. Biochem Biophys Res Commun. 2009; 381:660–5.23. Stoltzman CA, Peterson CW, Breen KT, Muoio DM, Billin AN, Ayer DE. Glucose sensing by MondoA:Mlx complexes: a role for hexokinases and direct regulation of thioredoxin-interacting protein expression. Proc Natl Acad Sci U S A. 2008; 105:6912–7.24. Kim DH, Kim JY, Yu BP, Chung HY. The activation of NFkappaB through Akt-induced FOXO1 phosphorylation during aging and its modulation by calorie restriction. Biogerontology. 2008; 9:33–47.25. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970; 227:680–5.26. Schulze PC, Yoshioka J, Takahashi T, He Z, King GL, Lee RT. Hyperglycemia promotes oxidative stress through inhibition of thioredoxin function by thioredoxin-interacting protein. J Biol Chem. 2004; 279:30369–74.27. Nivet-Antoine V, Cottart CH, Lemarechal H, Vamy M, Margaill I, Beaudeux JL, et al. trans-Resveratrol downregulates Txnip overexpression occurring during liver ischemia-reperfusion. Biochimie. 2010; 92:1766–71.28. Chutkow WA, Patwari P, Yoshioka J, Lee RT. Thioredoxininteracting protein (Txnip) is a critical regulator of hepatic glucose production. J Biol Chem. 2008; 283:2397–406.29. Boaru SG, Borkham-Kamphorst E, Tihaa L, Haas U, Weiskirchen R. Expression analysis of inflammasomes in experimental models of inflammatory and fibrotic liver disease. J Inflamm (Lond). 2012; 9:49.30. Calabuig-Navarro V, Yamauchi J, Lee S, Zhang T, Liu YZ, Sadlek K, et al. Forkhead box O6 (FoxO6) depletion attenuates hepatic gluconeogenesis and protects against fat-induced glucose disorder in mice. J Biol Chem. 2015; 290:15581–94.31. Al-Mubarak B, Soriano FX, Hardingham GE. Synaptic NMDAR activity suppresses FOXO1 expression via a cis-acting FOXO binding site: FOXO1 is a FOXO target gene. Channels (Austin). 2009; 3:233–8.32. Xuan Z, Zhang MQ. From worm to human: bioinformatics approaches to identify FOXO target genes. Mech Ageing Dev. 2005; 126:209–15.33. Yamaguchi F, Hirata Y, Akram H, Kamitori K, Dong Y, Sui L, et al. FOXO/TXNIP pathway is involved in the suppression of hepatocellular carcinoma growth by glutamate antagonist MK-801. BMC Cancer. 2013; 13:468.34. Kim DH, Zhang T, Lee S, Calabuig-Navarro V, Yamauchi J, Piccirillo A, et al. FoxO6 integrates insulin signaling with MTP for regulating VLDL production in the liver. Endocrinology. 2014; 155:1255–67.35. Goossens GH, Blaak EE, Theunissen R, Duijvestijn AM, Clement K, Tervaert JW, et al. Expression of NLRP3 inflammasome and T cell population markers in adipose tissue are associated with insulin resistance and impaired glucose metabolism in humans. Mol Immunol. 2012; 50:142–9.36. Csak T, Ganz M, Pespisa J, Kodys K, Dolganiuc A, Szabo G. Fatty acid and endotoxin activate inflammasomes in mouse hepatocytes that release danger signals to stimulate immune cells. Hepatology. 2011; 54:133–44.37. Wen H, Gris D, Lei Y, Jha S, Zhang L, Huang MT, et al. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat Immunol. 2011; 12:408–15.38. Guha M, Mackman N. The phosphatidylinositol 3-kinaseAkt pathway limits lipopolysaccharide activation of signaling pathways and expression of inflammatory mediators in human monocytic cells. J Biol Chem. 2002; 277:32124–32.39. Molnarfi N, Gruaz L, Dayer JM, Burger D. Opposite regulation of IL-1beta and secreted IL-1 receptor antagonist production by phosphatidylinositide-3 kinases in human monocytes activated by lipopolysaccharides or contact with T cells. J Immunol. 2007; 178:446–54.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Ethanol Augments Monosodium Urate-Induced NLRP3 Inflammasome Activation via Regulation of AhR and TXNIP in Human Macrophages
- Loganin Prevents Hepatic Steatosis by Blocking NLRP3 Inflammasome Activation
- The Mechanism of the NLRP3 Inflammasome Activation and Pathogenic Implication in the Pathogenesis of Gout
- NLRP3 Inflammasome and Host Protection against Bacterial Infection
- Extracellular Acidification Augments NLRP3-Mediated Inflammasome Signaling in Macrophages
